

Identification of amino acids in the N-terminal domain of atypical methanogenic-type Seryl-tRNA synthetase critical for tRNA recognition
- PMID: 19734148
- PMCID: PMC2781618
- DOI: 10.1074/jbc.M109.044099
Identification of amino acids in the N-terminal domain of atypical methanogenic-type Seryl-tRNA synthetase critical for tRNA recognition
Abstract
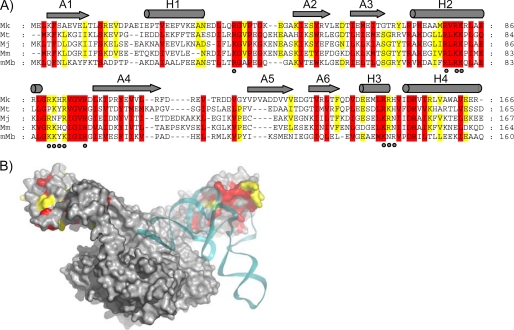
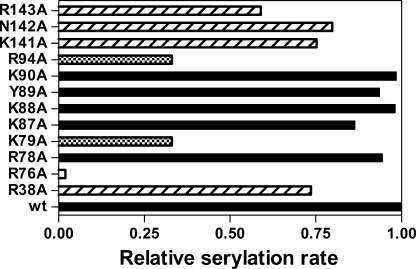
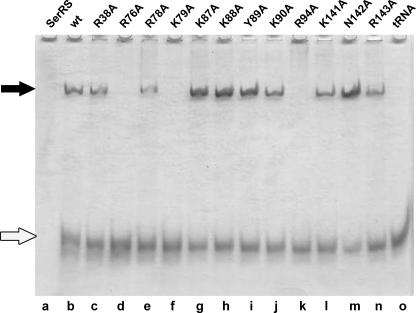
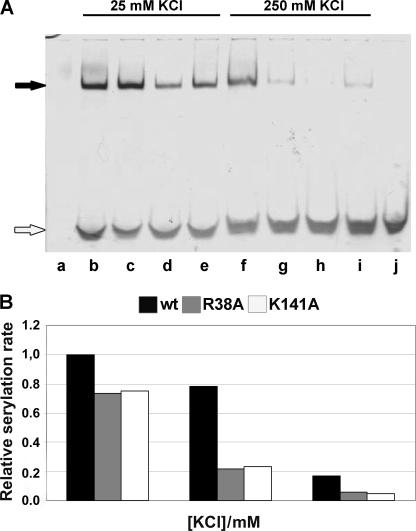
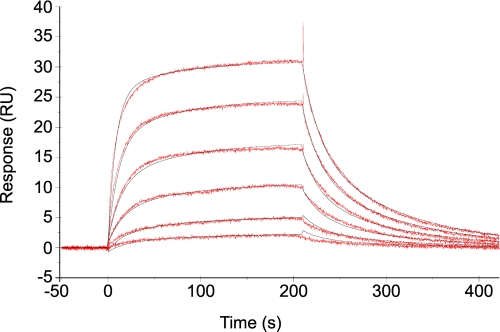
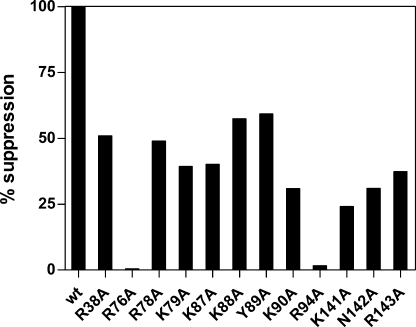
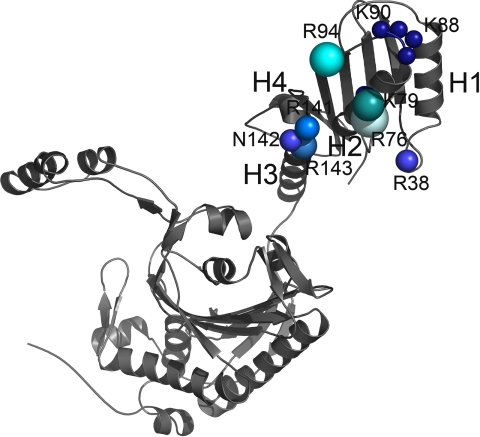
Seryl-tRNA synthetase (SerRS) from methanogenic archaeon Methanosarcina barkeri, contains an idiosyncratic N-terminal domain, composed of an antiparallel beta-sheet capped by a helical bundle, connected to the catalytic core by a short linker peptide. It is very different from the coiled-coil tRNA binding domain in bacterial-type SerRS. Because the crystal structure of the methanogenic-type SerRSxtRNA complex has not been obtained, a docking model was produced, which indicated that highly conserved helices H2 and H3 of the N-terminal domain may be important for recognition of the extra arm of tRNA(Ser). Based on structural information and the docking model, we have mutated various positions within the N-terminal region and probed their involvement in tRNA binding and serylation. Total loss of activity and inability of the R76A variant to form the complex with cognate tRNA identifies Arg(76) located in helix H2 as a crucial tRNA-interacting residue. Alteration of Lys(79) positioned in helix H2 and Arg(94) in the loop between helix H2 and beta-strand A4 have a pronounced effect on SerRSxtRNA(Ser) complex formation and dissociation constants (K(D)) determined by surface plasmon resonance. The replacement of residues Arg(38) (located in the loop between helix H1 and beta-strand A2), Lys(141) and Asn(142) (from H3), and Arg(143) (between H3 and H4) moderately affect both the serylation activity and the K(D) values. Furthermore, we have obtained a striking correlation between these results and in vivo effects of these mutations by quantifying the efficiency of suppression of bacterial amber mutations, after coexpression of the genes for M. barkeri suppressor tRNA(Ser) and a set of mMbSerRS variants in Escherichia coli.
Figures
References
-
- Ibba M., Soll D. (2000) Annu. Rev. Biochem. 69, 617–650 - PubMed
-
- Eriani G., Delarue M., Poch O., Gangloff J., Moras D. (1990) Nature 347, 203–206 - PubMed
-
- Cusack S., Berthet-Colominas C., Härtlein M., Nassar N., Leberman R. (1990) Nature 347, 249–255 - PubMed
-
- Ibba M., Morgan S., Curnow A. W., Pridmore D. R., Vothknecht U. C., Gardner W., Lin W., Woese C. R., Söll D. (1997) Science 278, 1119–1122 - PubMed
-
- Zhang C. M., Perona J. J., Ryu K., Francklyn C., Hou Y. M. (2006) J. Mol. Biol. 361, 300–311 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
