

Quantitating T cell cross-reactivity for unrelated peptide antigens
- PMID: 19734234
- PMCID: PMC2762195
- DOI: 10.4049/jimmunol.0901607
Quantitating T cell cross-reactivity for unrelated peptide antigens
Abstract
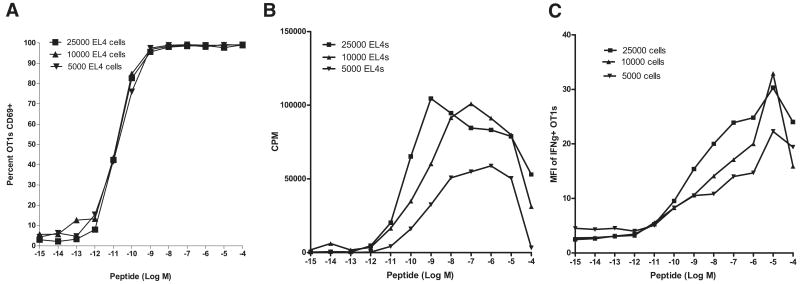
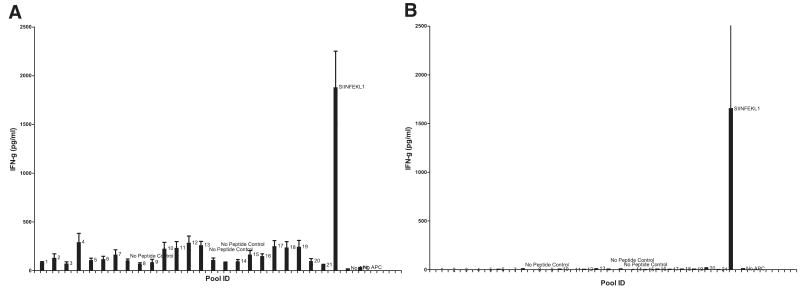
Quantitating the frequency of T cell cross-reactivity to unrelated peptides is essential to understanding T cell responses in infectious and autoimmune diseases. Here we used 15 mouse or human CD8+ T cell clones (11 antiviral, 4 anti-self) in conjunction with a large library of defined synthetic peptides to examine nearly 30,000 TCR-peptide MHC class I interactions for cross-reactions. We identified a single cross-reaction consisting of an anti-self TCR recognizing a poxvirus peptide at relatively low sensitivity. We failed to identify any cross-reactions between the synthetic peptides in the panel and polyclonal CD8+ T cells raised to viral or alloantigens. These findings provide the best estimate to date of the frequency of T cell cross-reactivity to unrelated peptides ( approximately 1/30,000), explaining why cross-reactions between unrelated pathogens are infrequently encountered and providing a critical parameter for understanding the scope of self-tolerance.
Conflict of interest statement
Figures




References
-
- Manning TC, Schlueter CJ, Brodnicki TC, Parke EA, Speir JA, Garcia KC, Teyton L, Wilson IA, Kranz DM. Alanine scanning mutagenesis of an αβ T cell receptor: mapping the energy of antigen recognition. Immunity. 1998;8:413–425. - PubMed
-
- Kan-Mitchell J, Bajcz M, Schaubert KL, Price DA, Brenchley JM, Asher TE, Douek DC, Ng HL, Yang OO, Rinaldo CR, Jr, et al. Degeneracy and repertoire of the human HIV-1 Gag p1777–85 CTL response. J Immunol. 2006;176:6690–6701. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
