

The rkp-1 cluster is required for secretion of Kdo homopolymeric capsular polysaccharide in Sinorhizobium meliloti strain Rm1021
- PMID: 19734304
- PMCID: PMC2772494
- DOI: 10.1128/JB.00466-09
The rkp-1 cluster is required for secretion of Kdo homopolymeric capsular polysaccharide in Sinorhizobium meliloti strain Rm1021
Abstract
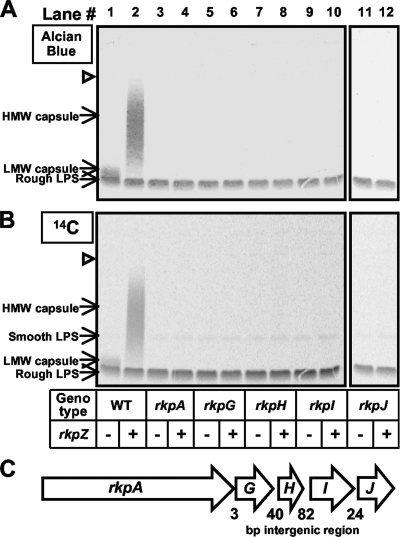
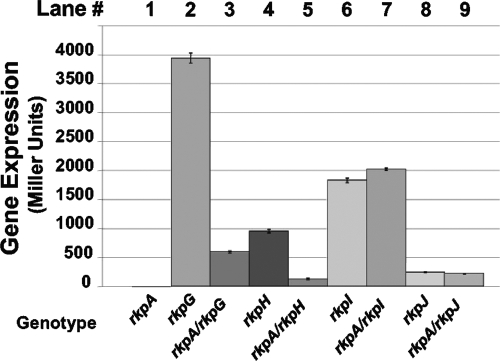
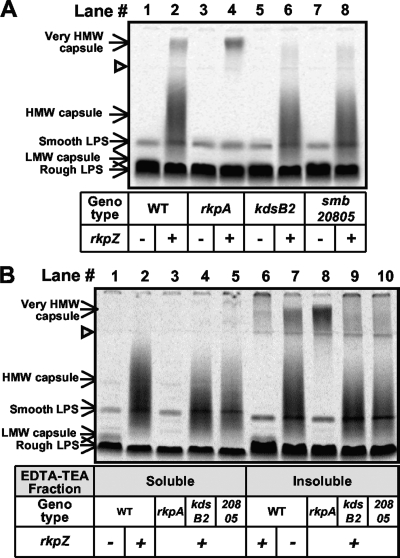
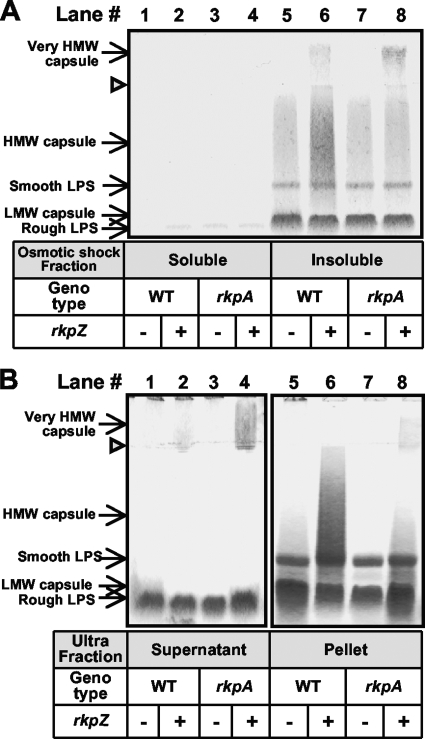
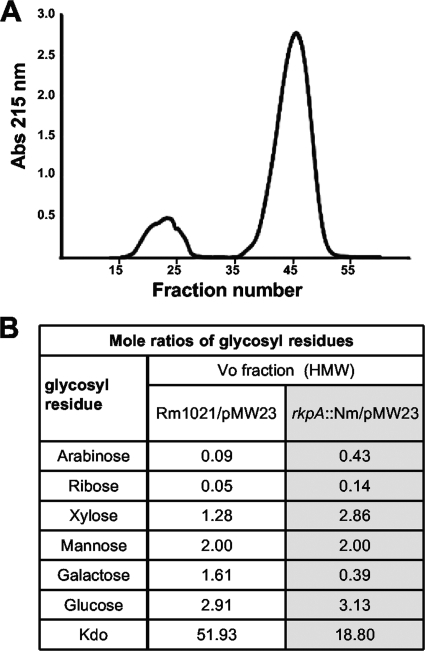
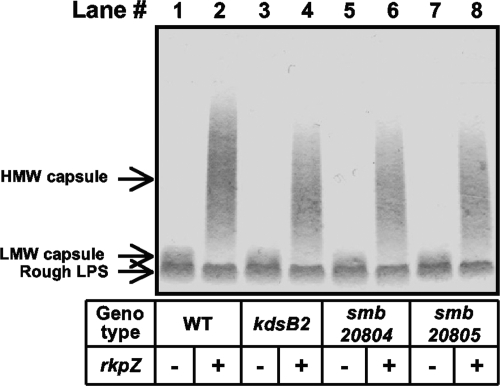
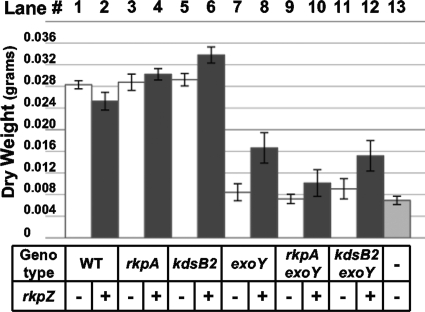
Under conditions of nitrogen stress, leguminous plants form symbioses with soil bacteria called rhizobia. This partnership results in the development of structures called root nodules, in which differentiated endosymbiotic bacteria reduce molecular dinitrogen for the host. The establishment of rhizobium-legume symbioses requires the bacterial synthesis of oligosaccharides, exopolysaccharides, and capsular polysaccharides. Previous studies suggested that the 3-deoxy-D-manno-oct-2-ulopyranosonic acid (Kdo) homopolymeric capsular polysaccharide produced by strain Sinorhizobium meliloti Rm1021 contributes to symbiosis with Medicago sativa under some conditions. However, a conclusive symbiotic role for this polysaccharide could not be determined due to a lack of mutants affecting its synthesis. In this study, we have further characterized the synthesis, secretion, and symbiotic function of the Kdo homopolymeric capsule. We showed that mutants lacking the enigmatic rkp-1 gene cluster fail to display the Kdo capsule on the cell surface but accumulate an intracellular polysaccharide of unusually high M(r). In addition, we have demonstrated that mutations in kdsB2, smb20804, and smb20805 affect the polymerization of the Kdo homopolymeric capsule. Our studies also suggest a role for the capsular polysaccharide in symbiosis. Previous reports have shown that the overexpression of rkpZ from strain Rm41 allows for the symbiosis of exoY mutants of Rm1021 that are unable to produce the exopolysaccharide succinoglycan. Our results demonstrate that mutations in the rkp-1 cluster prevent this phenotypic suppression of exoY mutants, although mutations in kdsB2, smb20804, and smb20805 have no effect.
Figures
Similar articles
-
Overproduction and increased molecular weight account for the symbiotic activity of the rkpZ-modified K polysaccharide from Sinorhizobium meliloti Rm1021.Glycobiology. 2006 Dec;16(12):1181-93. doi: 10.1093/glycob/cwl042. Epub 2006 Sep 6. Glycobiology. 2006. PMID: 16957092
-
Sinorhizobium medicae WSM419 Genes That Improve Symbiosis between Sinorhizobium meliloti Rm1021 and Medicago truncatula Jemalong A17 and in Other Symbiosis Systems.Appl Environ Microbiol. 2021 Jul 13;87(15):e0300420. doi: 10.1128/AEM.03004-20. Epub 2021 Jul 13. Appl Environ Microbiol. 2021. PMID: 33990306 Free PMC article.
-
Genetic analysis of the rkp-3 gene region in Sinorhizobium meliloti 41: rkpY directs capsular polysaccharide synthesis to KR5 antigen production.Mol Plant Microbe Interact. 2009 Nov;22(11):1422-30. doi: 10.1094/MPMI-22-11-1422. Mol Plant Microbe Interact. 2009. PMID: 19810811
-
Rhizobium meliloti exopolysaccharides: synthesis and symbiotic function.Gene. 1996 Nov 7;179(1):141-6. doi: 10.1016/s0378-1119(96)00322-8. Gene. 1996. PMID: 8955640 Review.
-
Polyamine biosynthesis and biological roles in rhizobia.FEMS Microbiol Lett. 2019 Apr 1;366(7):fnz084. doi: 10.1093/femsle/fnz084. FEMS Microbiol Lett. 2019. PMID: 31062028 Review.
Cited by
-
Role for Rhizobium rhizogenes K84 cell envelope polysaccharides in surface interactions.Appl Environ Microbiol. 2012 Mar;78(6):1644-51. doi: 10.1128/AEM.07117-11. Epub 2011 Dec 30. Appl Environ Microbiol. 2012. PMID: 22210213 Free PMC article.
-
Genome-Wide Sensitivity Analysis of the Microsymbiont Sinorhizobium meliloti to Symbiotically Important, Defensin-Like Host Peptides.mBio. 2017 Aug 1;8(4):e01060-17. doi: 10.1128/mBio.01060-17. mBio. 2017. PMID: 28765224 Free PMC article.
-
ABC transporters involved in export of cell surface glycoconjugates.Microbiol Mol Biol Rev. 2010 Sep;74(3):341-62. doi: 10.1128/MMBR.00009-10. Microbiol Mol Biol Rev. 2010. PMID: 20805402 Free PMC article. Review.
-
Bacterial Molecular Signals in the Sinorhizobium fredii-Soybean Symbiosis.Int J Mol Sci. 2016 May 18;17(5):755. doi: 10.3390/ijms17050755. Int J Mol Sci. 2016. PMID: 27213334 Free PMC article.
-
A Clostridium difficile Cell Wall Glycopolymer Locus Influences Bacterial Shape, Polysaccharide Production and Virulence.PLoS Pathog. 2016 Oct 14;12(10):e1005946. doi: 10.1371/journal.ppat.1005946. eCollection 2016 Oct. PLoS Pathog. 2016. PMID: 27741317 Free PMC article.
References
-
- Barnett, M. J., R. F. Fisher, T. Jones, C. Komp, A. P. Abola, F. Barloy-Hubler, L. Bowser, D. Capela, F. Galibert, J. Gouzy, M. Gurjal, A. Hong, L. Huizar, R. W. Hyman, D. Kahn, M. L. Kahn, S. Kalman, D. H. Keating, C. Palm, M. C. Peck, R. Surzycki, D. H. Wells, K. C. Yeh, R. W. Davis, N. A. Federspiel, and S. R. Long. 2001. Nucleotide sequence and predicted functions of the entire Sinorhizobium meliloti pSymA megaplasmid. Proc. Natl. Acad. Sci. USA 98:9883-9888. - PMC - PubMed
-
- Barnett, M. J., V. Oke, and S. R. Long. 2000. New genetic tools for use in the Rhizobiaceae and other bacteria. BioTechniques 29:240-245. - PubMed
-
- Becker, A., N. Fraysse, and L. Sharypova. 2005. Recent advances in studies on structure and symbiosis-related function of rhizobial K-antigens and lipopolysaccharides. Mol. Plant-Microbe Interact. 18:899-905. - PubMed
-
- Beringer, J. E. 1974. R factor transfer in Rhizobium leguminosarum. J. Gen. Microbiol. 84:188-198. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
