

Neuron-glia communication via EphA4/ephrin-A3 modulates LTP through glial glutamate transport
- PMID: 19734893
- PMCID: PMC3922060
- DOI: 10.1038/nn.2394
Neuron-glia communication via EphA4/ephrin-A3 modulates LTP through glial glutamate transport
Abstract
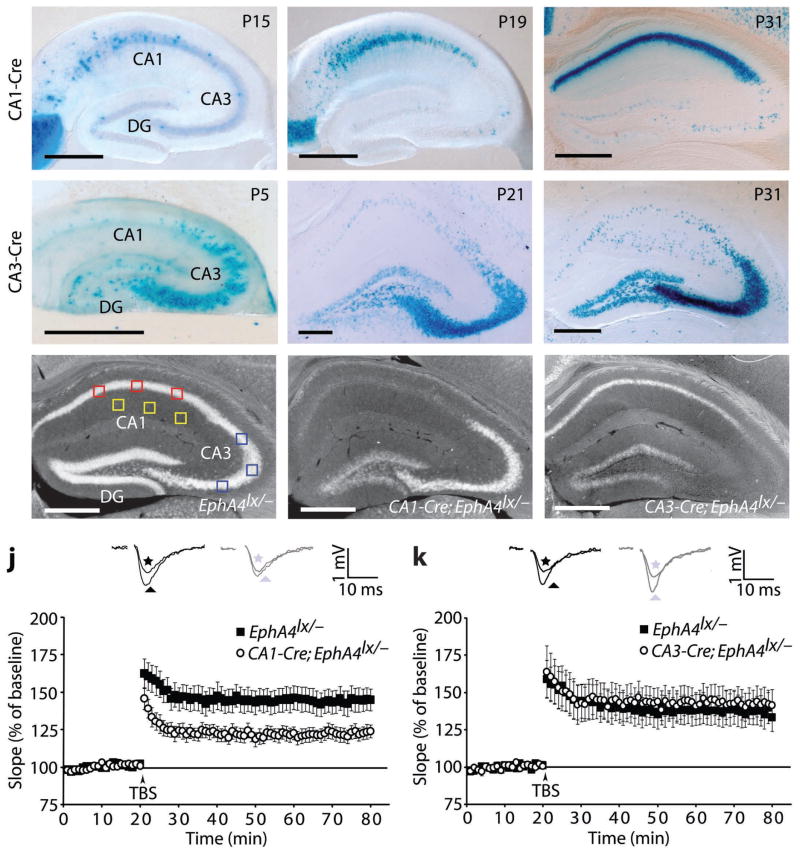
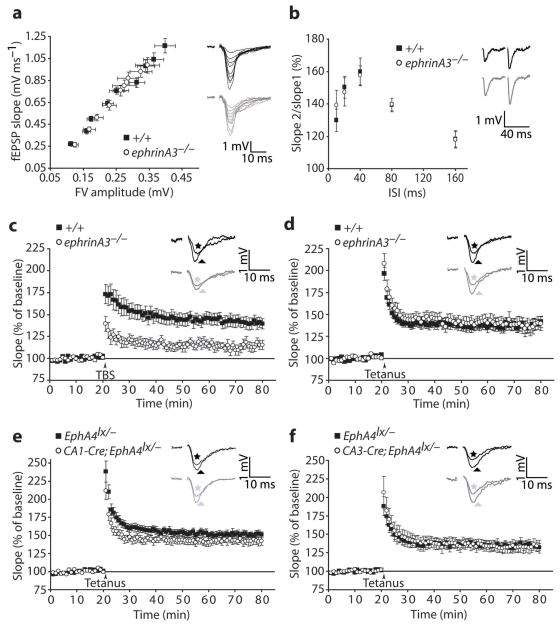
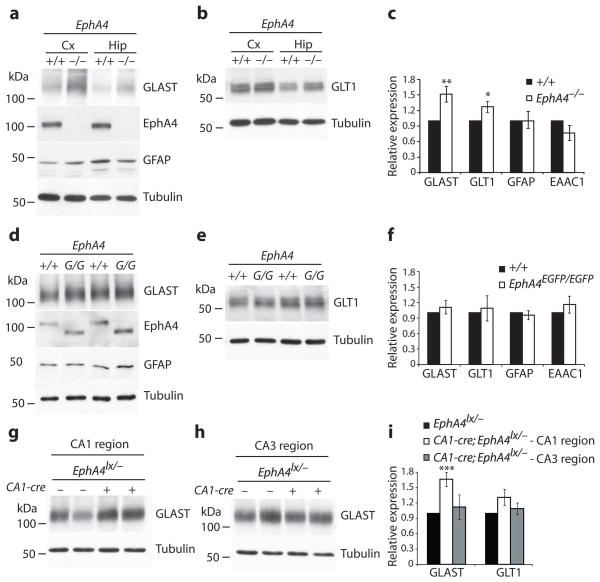
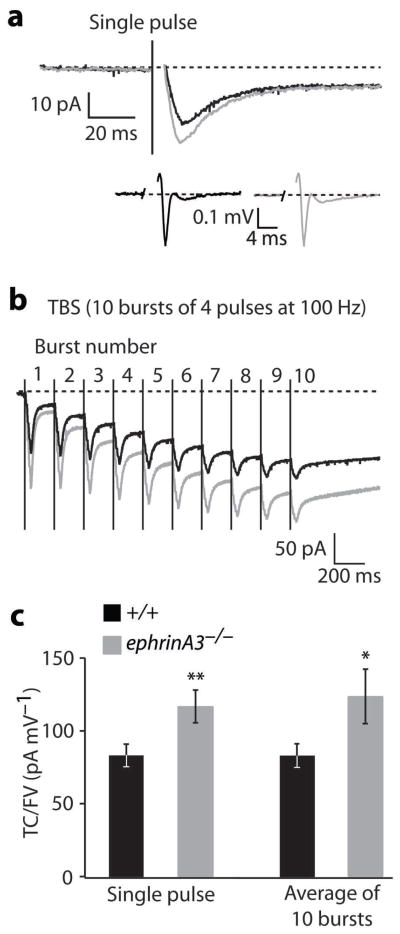
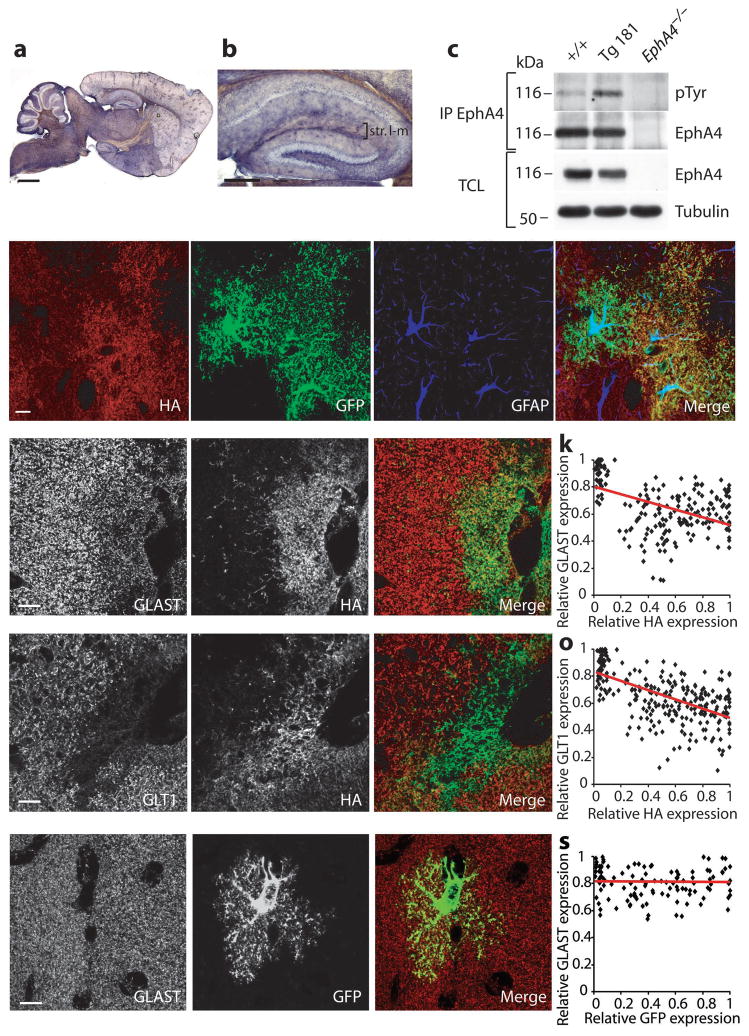
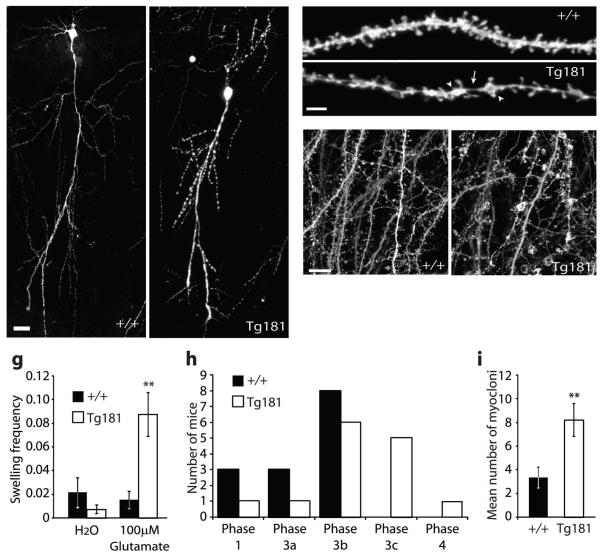
Astrocytes are critical participants in synapse development and function, but their role in synaptic plasticity is unclear. Eph receptors and their ephrin ligands have been suggested to regulate neuron-glia interactions, and EphA4-mediated ephrin reverse signaling is required for synaptic plasticity in the hippocampus. Here we show that long-term potentiation (LTP) at the CA3-CA1 synapse is modulated by EphA4 in the postsynaptic CA1 cell and by ephrin-A3, a ligand of EphA4 that is found in astrocytes. Lack of EphA4 increased the abundance of glial glutamate transporters, and ephrin-A3 modulated transporter currents in astrocytes. Pharmacological inhibition of glial glutamate transporters rescued the LTP defects in EphA4 (Epha4) and ephrin-A3 (Efna3) mutant mice. Transgenic overexpression of ephrin-A3 in astrocytes reduces glutamate transporter levels and produces focal dendritic swellings possibly caused by glutamate excitotoxicity. These results suggest that EphA4/ephrin-A3 signaling is a critical mechanism for astrocytes to regulate synaptic function and plasticity.
Figures

References
-
- Allen NJ, Barres BA. Signaling between glia and neurons: focus on synaptic plasticity. Current opinion in neurobiology. 2005;15:542–548. - PubMed
-
- Barres BA. The mystery and magic of glia: a perspective on their roles in health and disease. Neuron. 2008;60:430–440. - PubMed
-
- Halassa MM, Fellin T, Haydon PG. The tripartite synapse: roles for gliotransmission in health and disease. Trends in molecular medicine. 2007;13:54–63. - PubMed
-
- Tzingounis AV, Wadiche JI. Glutamate transporters: confining runaway excitation by shaping synaptic transmission. Nature reviews. 2007;8:935–947. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
