

Differential expression of membrane conductances underlies spontaneous event initiation by rostral midline neurons in the embryonic mouse hindbrain
- PMID: 19736299
- PMCID: PMC2790250
- DOI: 10.1113/jphysiol.2009.180091
Differential expression of membrane conductances underlies spontaneous event initiation by rostral midline neurons in the embryonic mouse hindbrain
Abstract
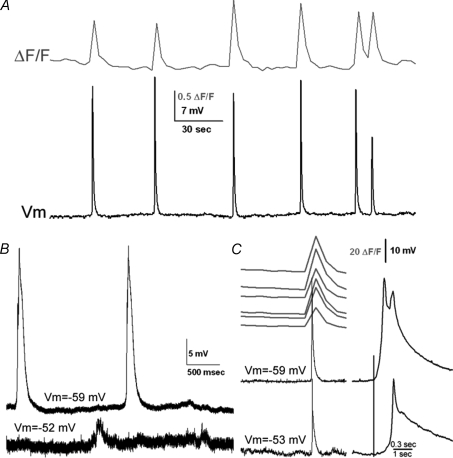
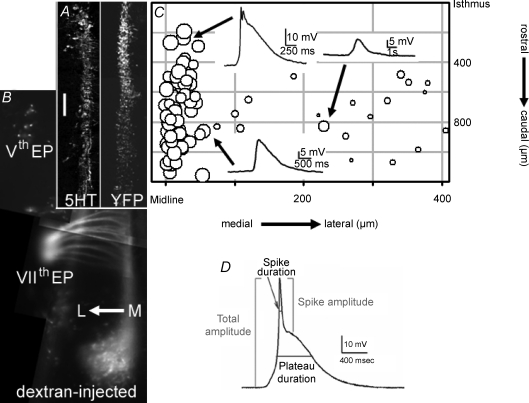
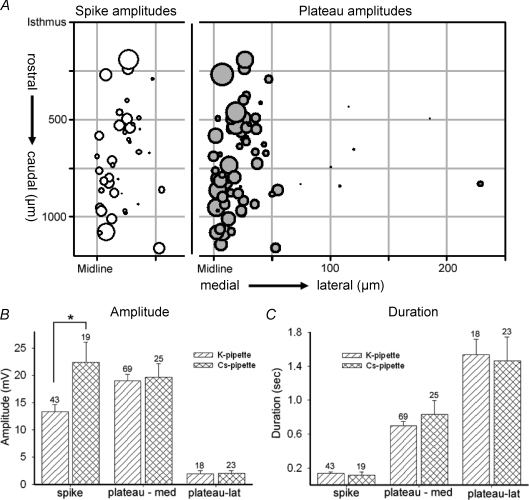
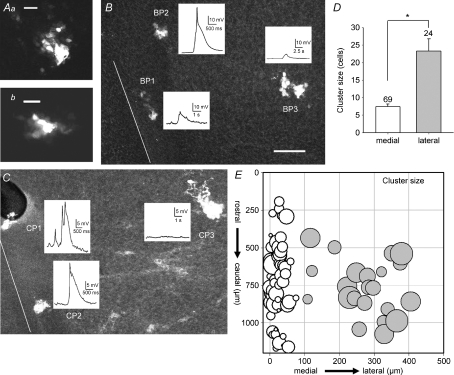
Spontaneous activity is expressed in many developing CNS structures and is crucial in correct network development. Previous work using [Ca(2+)](i) imaging showed that in the embryonic mouse hindbrain spontaneous activity is initiated by a driver population, the serotonergic neurons of the nascent raphe. Serotonergic neurons derived from former rhombomere 2 drive 90% of all hindbrain events at E11.5. We now demonstrate that the electrical correlate of individual events is a spontaneous depolarization, which originates at the rostral midline and drives events laterally. Midline events have both a rapid spike and a large plateau component, while events in lateral tissue comprise only a smaller amplitude plateau. Lateral cells have a large resting conductance and are highly coupled via neurobiotin-permeant gap junctions, while midline cells are significantly less gap junction-coupled and uniquely express a T-type Ca(2+) channel. We propose that the combination of low resting conductance and expression of T-type Ca(2+) current is permissive for midline neurons to acquire the initiator or driver phenotype, while cells without these features cannot drive activity. This demonstrates that expression of specific conductances contributes to the ability to drive spontaneous activity in a developing network.
Figures
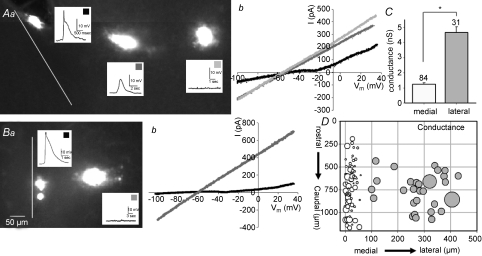
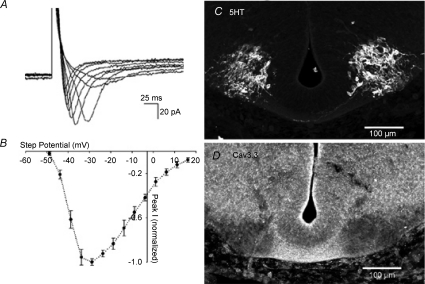
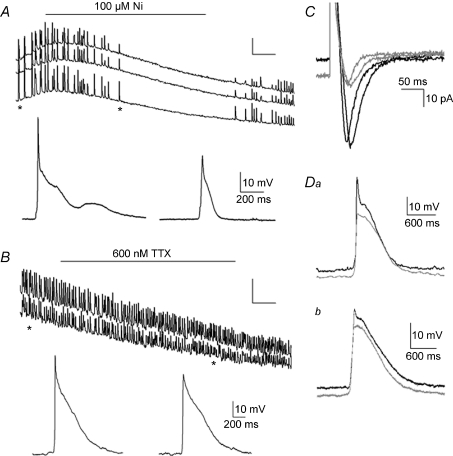
Comment in
-
Insights into medio-lateral signalling in the developing mouse hindbrain: properties of midline drivers of network activity.J Physiol. 2009 Nov 1;587(Pt 21):5007. doi: 10.1113/jphysiol.2009.181347. J Physiol. 2009. PMID: 19880874 Free PMC article. No abstract available.
References
-
- Aghajanian GK, Vandermaelen CP. Intracellular recordings from serotonergic dorsal raphe neurons: pacemaker potentials and the effect of LSD. Brain Res. 1982;238:463–469. - PubMed
-
- Bayliss DA, Li YW, Talley EM. Effects of serotonin on caudal raphe neurons: activation of an inwardly rectifying potassium conductance. J Neurophysiol. 1997a;77:1349–1361. - PubMed
-
- Bayliss DA, Li YW, Talley EM. Effects of serotonin on caudel raphe neurons: inhibition of N- and P/Q-type calcium channels and the afterhyperpolarization. J Neurophysiol. 1997b;77:1362–1374. - PubMed
-
- Ben-Ari Y. Developing networks play a similar melody. Trends Neurosci. 2001;24:353–360. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous