

Disruption of the pollen-expressed FERONIA homologs ANXUR1 and ANXUR2 triggers pollen tube discharge
- PMID: 19736323
- PMCID: PMC2739144
- DOI: 10.1242/dev.040071
Disruption of the pollen-expressed FERONIA homologs ANXUR1 and ANXUR2 triggers pollen tube discharge
Abstract
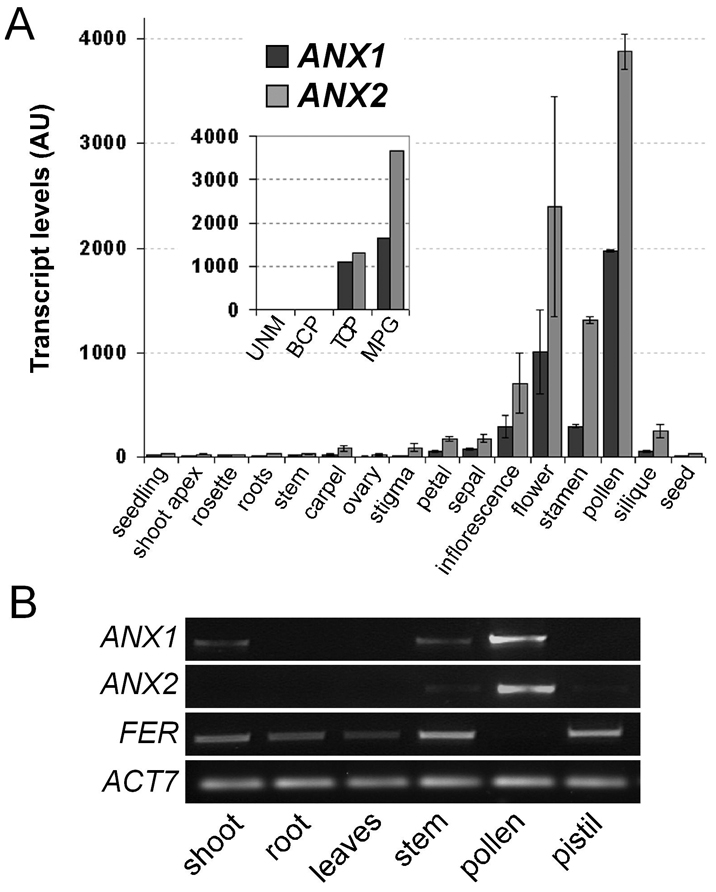
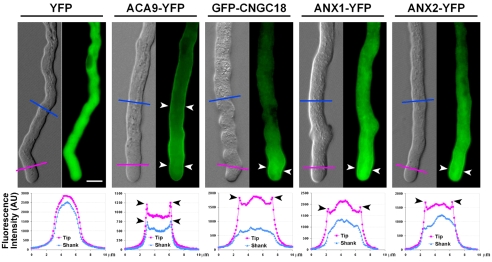
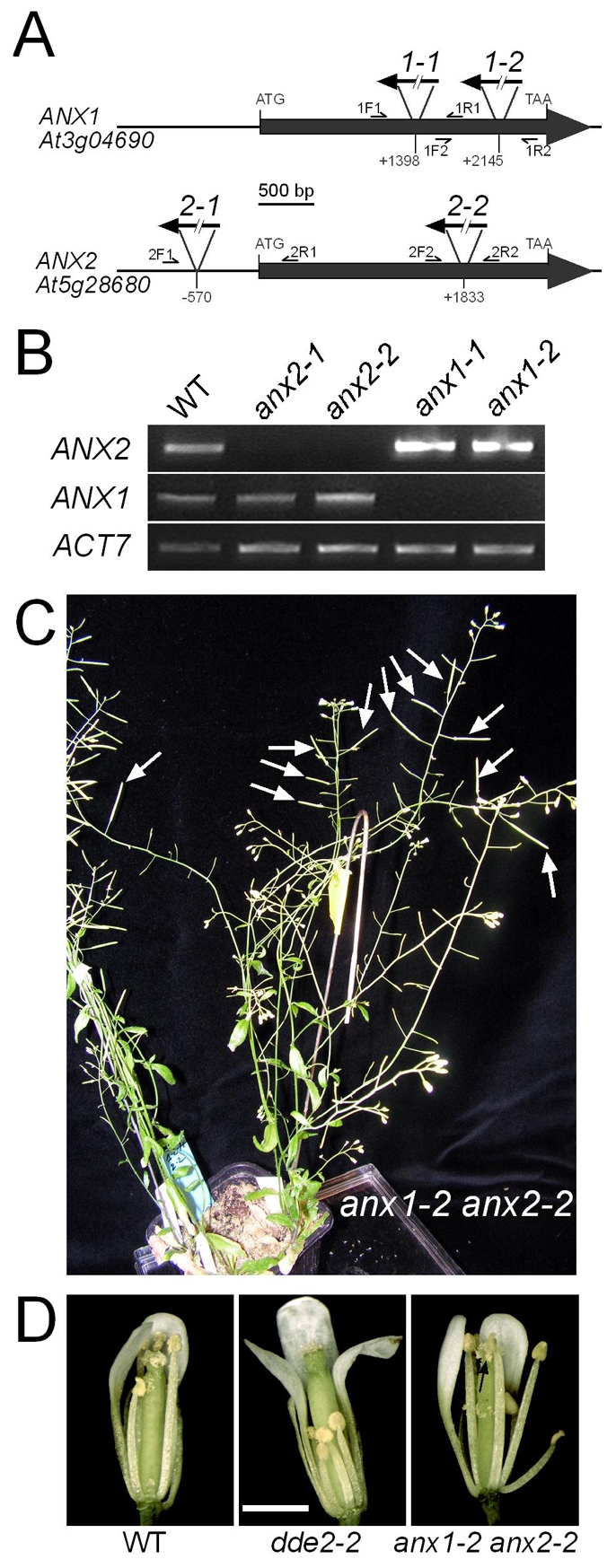
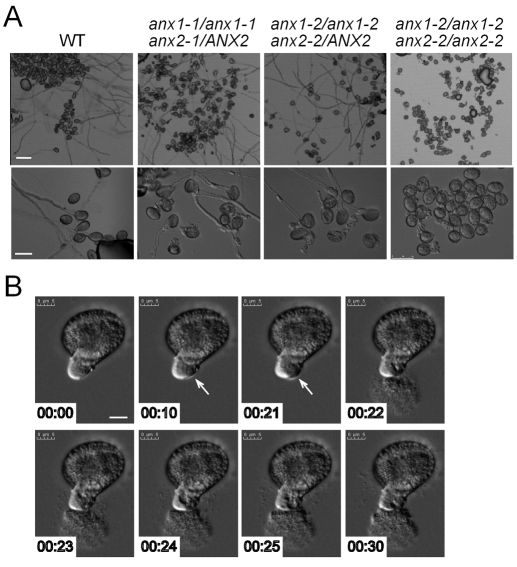
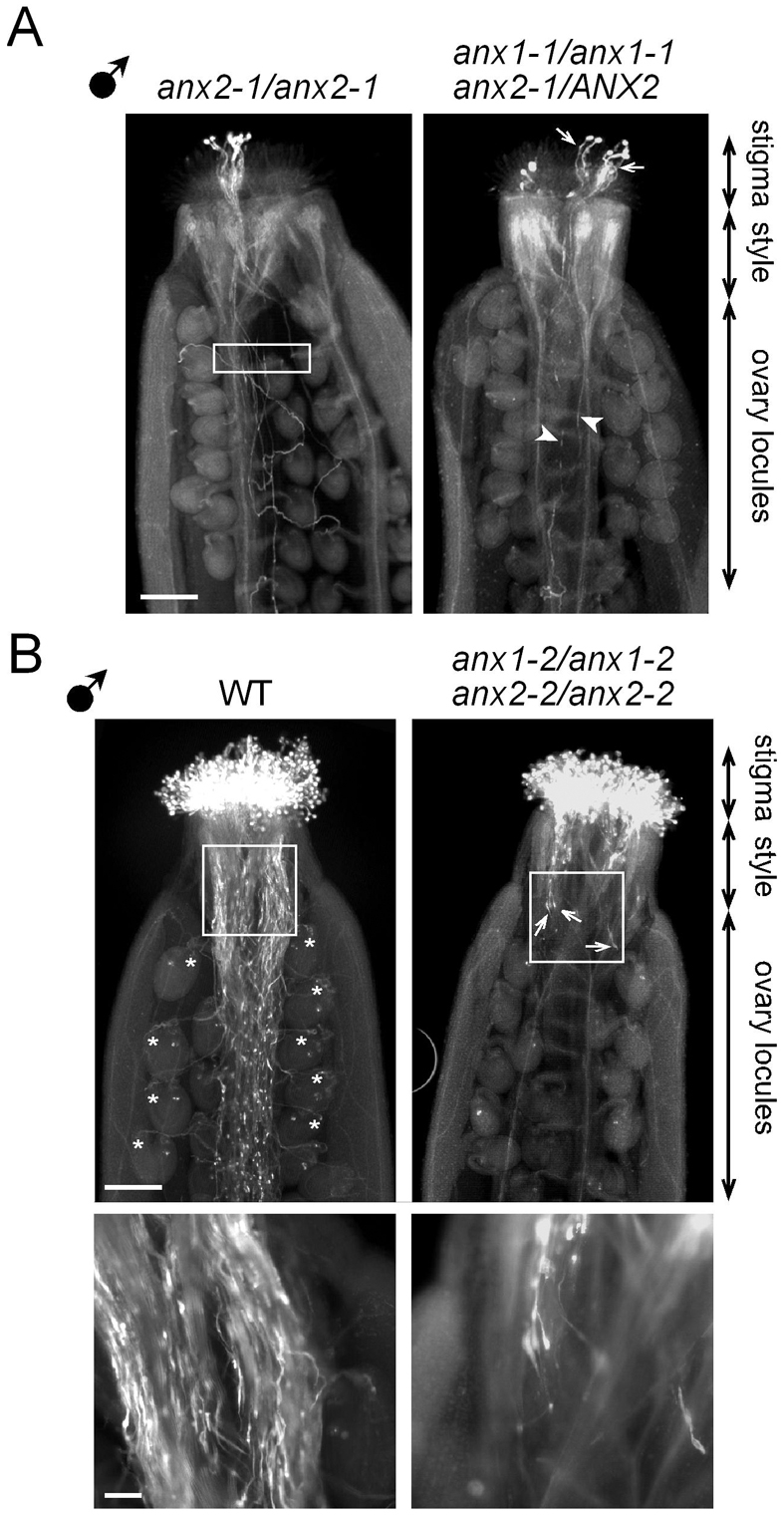
The precise delivery of male to female gametes during reproduction in eukaryotes requires complex signal exchanges and a flawless communication between male and female tissues. In angiosperms, molecular mechanisms have recently been revealed that are crucial for the dialog between male (pollen tube) and female gametophytes required for successful sperm delivery. When pollen tubes reach the female gametophyte, they arrest growth, burst and discharge their sperm cells. These processes are under the control of the female gametophyte via the receptor-like serine-threonine kinase (RLK) FERONIA (FER). However, the male signaling components that control the sperm delivery remain elusive. Here, we show that ANXUR1 and ANXUR2 (ANX1, ANX2), which encode the closest homologs of the FER-RLK in Arabidopsis, are preferentially expressed in pollen. Moreover, ANX1-YFP and ANX2-YFP fusion proteins display polar localization to the plasma membrane at the tip of the pollen tube. Finally, genetic analyses demonstrate that ANX1 and ANX2 function redundantly to control the timing of pollen tube discharge as anx1 anx2 double-mutant pollen tubes cease their growth and burst in vitro and fail to reach the female gametophytes in vivo. We propose that ANX-RLKs constitutively inhibit pollen tube rupture and sperm discharge at the tip of growing pollen tubes to sustain their growth within maternal tissues until they reach the female gametophytes. Upon arrival, the female FER-dependent signaling cascade is activated to mediate pollen tube reception and fertilization, while male ANX-dependent signaling is deactivated, enabling the pollen tube to rupture and deliver its sperm cells to effect fertilization.
Figures
References
-
- Alonso, J. M., Stepanova, A. N., Leisse, T. J., Kim, C. J., Chen, H., Shinn, P., Stevenson, D. K., Zimmerman, J., Barajas, P., Cheuk, R. et al. (2003). Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 301, 653-657. - PubMed
-
- Berger, F., Hamamura, Y., Ingouff, M. and Higashiyama, T. (2008). Double fertilization-caught in the act. Trends Plant Sci. 13, 437-443. - PubMed
-
- Boavida, L. C. and McCormick, S. (2007). Temperature as a determinant factor for increased and reproducible in vitro pollen germination in Arabidopsis thaliana. Plant J. 52, 570-582. - PubMed
-
- Boavida, L. C., Vieira, A. M., Becker, J. D. and Feijo, J. A. (2005). Gametophyte interaction and sexual reproduction: how plants make a zygote. Int. J. Dev. Biol. 49, 615-632. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
