

Enhancement of tumour-specific immune responses in vivo by 'MHC loading-enhancer' (MLE)
- PMID: 19738910
- PMCID: PMC2735034
- DOI: 10.1371/journal.pone.0006811
Enhancement of tumour-specific immune responses in vivo by 'MHC loading-enhancer' (MLE)
Abstract
Background: Class II MHC molecules (MHC II) are cell surface receptors displaying short protein fragments for the surveillance by CD4+ T cells. Antigens therefore have to be loaded onto this receptor in order to induce productive immune responses. On the cell surface, most MHC II molecules are either occupied by ligands or their binding cleft has been blocked by the acquisition of a non-receptive state. Direct loading with antigens, as required during peptide vaccinations, is therefore hindered.
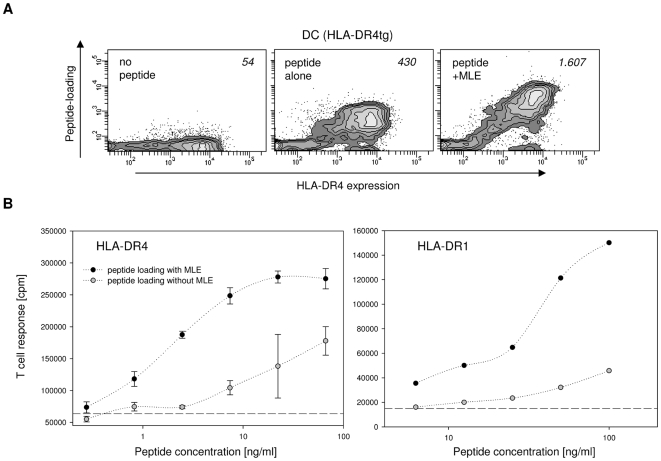
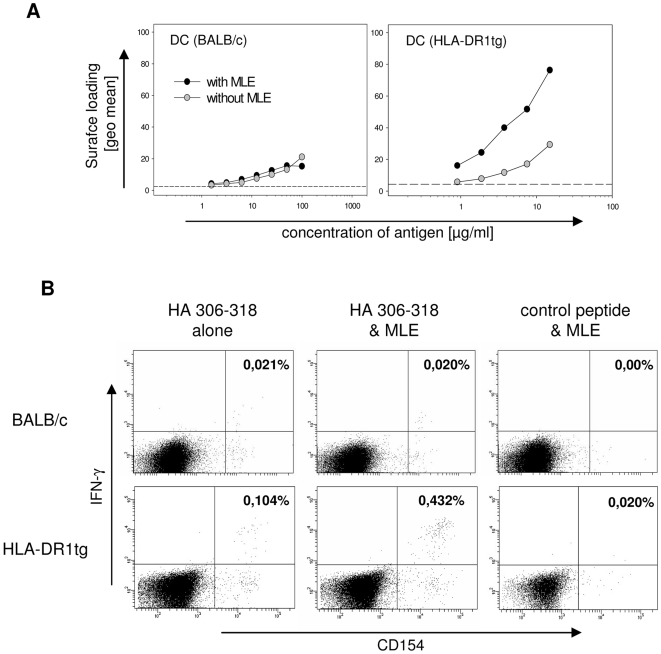
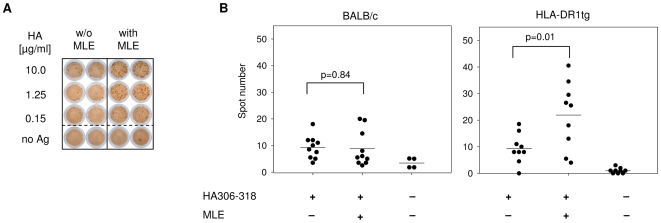
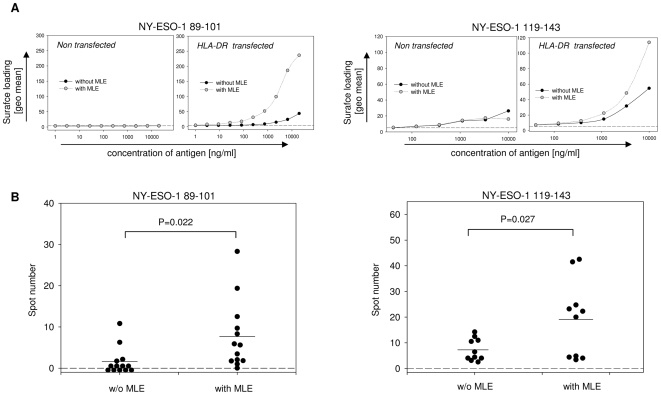
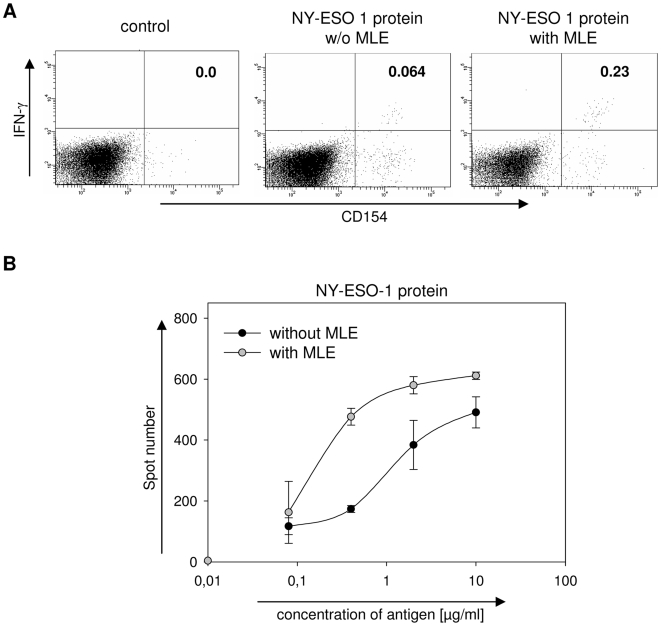
Principal findings: Here we show, that the in vivo response of CD4+ T cells can be improved, when the antigens are administered together with 'MHC-loading enhancer' (MLE). MLE are small catalytic compounds able to open up the MHC binding site by triggering ligand-release and stabilizing the receptive state. Their enhancing effect on the immune response was demonstrated here with an antigen from the influenza virus and tumour associated antigens (TAA) derived from the NY-ESO-1 protein. The application of these antigens in combination with adamantane ethanol (AdEtOH), an MLE compound active on human HLA-DR molecules, significantly increased the frequency of antigen-specific CD4+ T cells in mice transgenic for the human MHC II molecule. Notably, the effect was evident only with the MLE-susceptible HLA-DR molecule and not with murine MHC II molecules non-susceptible for the catalytic effect of the MLE.
Conclusion: MLE can specifically increase the potency of a vaccine by facilitating the efficient transfer of the antigen onto the MHC molecule. They may therefore open a new way to improve vaccination efficacy and tumour-immunotherapy.
Conflict of interest statement
Figures
Similar articles
-
A cyclic peptide accelerates the loading of peptide antigens in major histocompatibility complex class II molecules.Biochem Biophys Res Commun. 2015 Jan 16;456(3):774-9. doi: 10.1016/j.bbrc.2014.12.047. Epub 2014 Dec 15. Biochem Biophys Res Commun. 2015. PMID: 25522880
-
Exosomes as a tumor vaccine: enhancing potency through direct loading of antigenic peptides.J Immunother. 2003 Sep-Oct;26(5):440-50. doi: 10.1097/00002371-200309000-00007. J Immunother. 2003. PMID: 12973033
-
A universal anti-cancer vaccine: Chimeric invariant chain potentiates the inhibition of melanoma progression and the improvement of survival.Int J Cancer. 2019 Feb 15;144(4):909-921. doi: 10.1002/ijc.31795. Epub 2018 Dec 5. Int J Cancer. 2019. PMID: 30106470
-
CD4+ T-cell activation for immunotherapy of malignancies using Ii-Key/MHC class II epitope hybrid vaccines.Vaccine. 2012 Apr 16;30(18):2805-10. doi: 10.1016/j.vaccine.2012.02.031. Epub 2012 Mar 3. Vaccine. 2012. PMID: 22386748 Review.
-
Turning tumor cells in situ into T-helper cell-stimulating, MHC class II tumor epitope-presenters: immuno-curing and immuno-consolidation.Cancer Treat Rev. 2004 May;30(3):281-90. doi: 10.1016/j.ctrv.2003.08.002. Cancer Treat Rev. 2004. PMID: 15059651 Review.
Cited by
-
An immunoinformatic approach to assessing the immunogenic capacity of alpha-neurotoxins in elapid snake venoms.Front Pharmacol. 2023 Apr 10;14:1143437. doi: 10.3389/fphar.2023.1143437. eCollection 2023. Front Pharmacol. 2023. PMID: 37153801 Free PMC article.
-
T cell responses to human platelet antigen-1a involve a unique form of indirect allorecognition.JCI Insight. 2016 Sep 8;1(14):e86558. doi: 10.1172/jci.insight.86558. JCI Insight. 2016. PMID: 27699233 Free PMC article.
-
Activating interactions of sulfanilamides with T cell receptors.Open J Immunol. 2013 Sep;3(3):139-157. doi: 10.4236/oji.2013.33019. Open J Immunol. 2013. PMID: 36172594 Free PMC article.
-
Development and application of reverse genetic technology for the influenza virus.Virus Genes. 2021 Apr;57(2):151-163. doi: 10.1007/s11262-020-01822-9. Epub 2021 Feb 2. Virus Genes. 2021. PMID: 33528730 Free PMC article. Review.
-
On Peptides and Altered Peptide Ligands: From Origin, Mode of Action and Design to Clinical Application (Immunotherapy).Int Arch Allergy Immunol. 2016;170(4):211-233. doi: 10.1159/000448756. Epub 2016 Sep 20. Int Arch Allergy Immunol. 2016. PMID: 27642756 Free PMC article. Review.
References
-
- Villa LL, Costa RL, Petta CA, Andrade RP, Ault KA, et al. Prophylactic quadrivalent human papillomavirus (types 6, 11, 16, and 18) L1 virus-like particle vaccine in young women: a randomised double-blind placebo-controlled multicentre phase II efficacy trial. Lancet Oncol. 2005;6:271–278. - PubMed
-
- Harper DM, Franco EL, Wheeler C, Ferris DG, Jenkins D, et al. Efficacy of a bivalent L1 virus-like particle vaccine in prevention of infection with human papillomavirus types 16 and 18 in young women: a randomised controlled trial. Lancet. 2004;364:1757–1765. - PubMed
-
- Scanlan MJ, Gure AO, Jungbluth AA, Old LJ, Chen Y-T. Cancer/testis antigens: an expanding family of targets for cancer immunotherapy. Immunol Rev. 2002;188:22–32. - PubMed
-
- Gnjatic S, Nishikawa H, Jungbluth AA, Gure AO, Ritter G, et al. NY-ESO-1: review of an immunogenic tumor antigen. Adv Cancer Res. 2006;95:1–30. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
