

Endothelial cell specific adhesion molecule (ESAM) localizes to platelet-platelet contacts and regulates thrombus formation in vivo
- PMID: 19740102
- PMCID: PMC4441405
- DOI: 10.1111/j.1538-7836.2009.03606.x
Endothelial cell specific adhesion molecule (ESAM) localizes to platelet-platelet contacts and regulates thrombus formation in vivo
Abstract
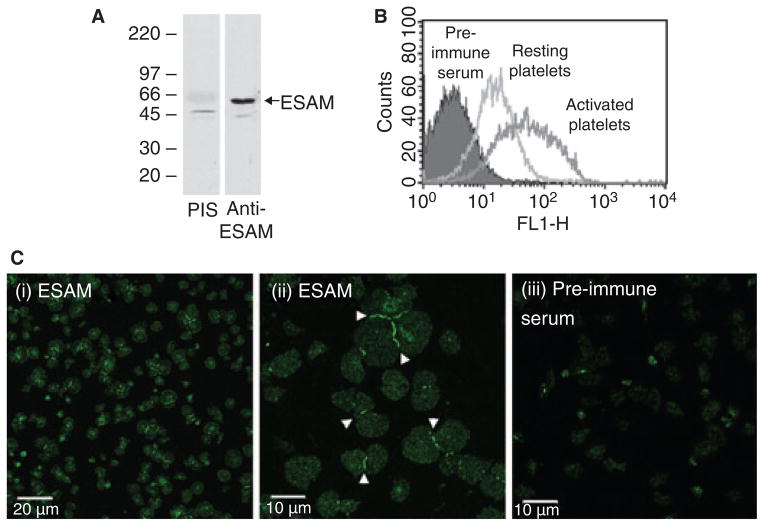
Background: In resting platelets, endothelial cell specific adhesion molecule (ESAM) is located in alpha granules, increasing its cell surface expression following platelet activation. However, the function of ESAM on platelets is unknown.
Objective: To determine whether ESAM has a role in thrombus formation.
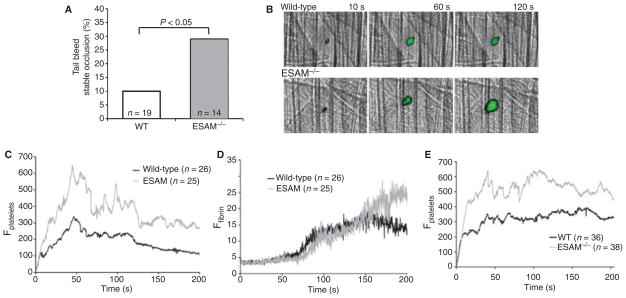
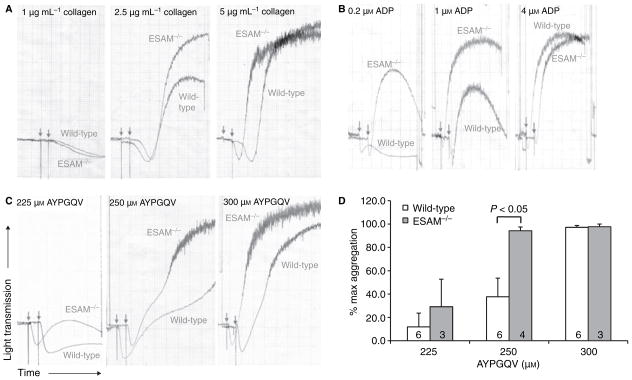
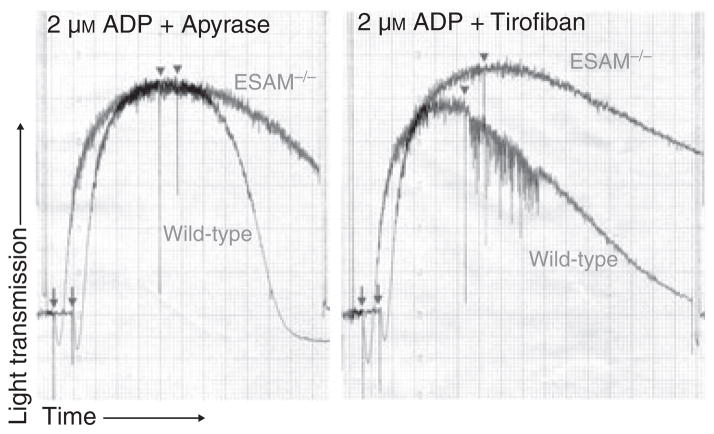
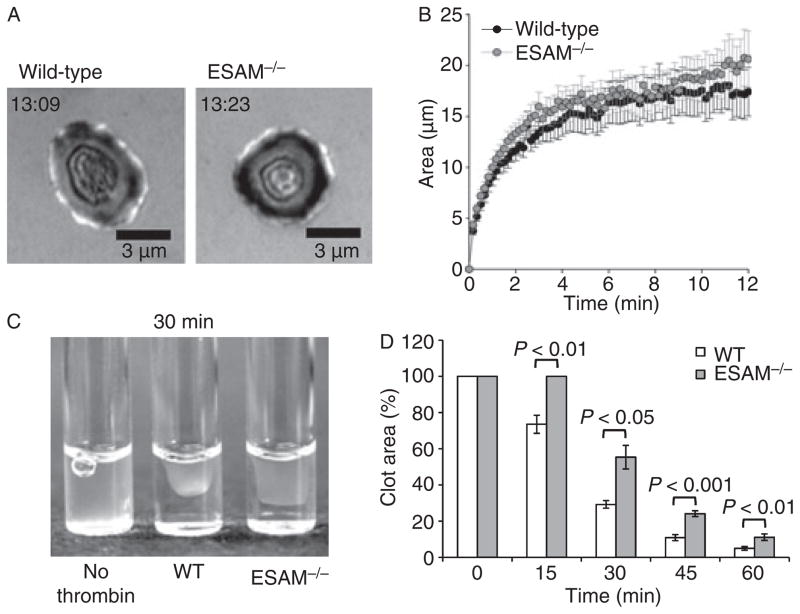
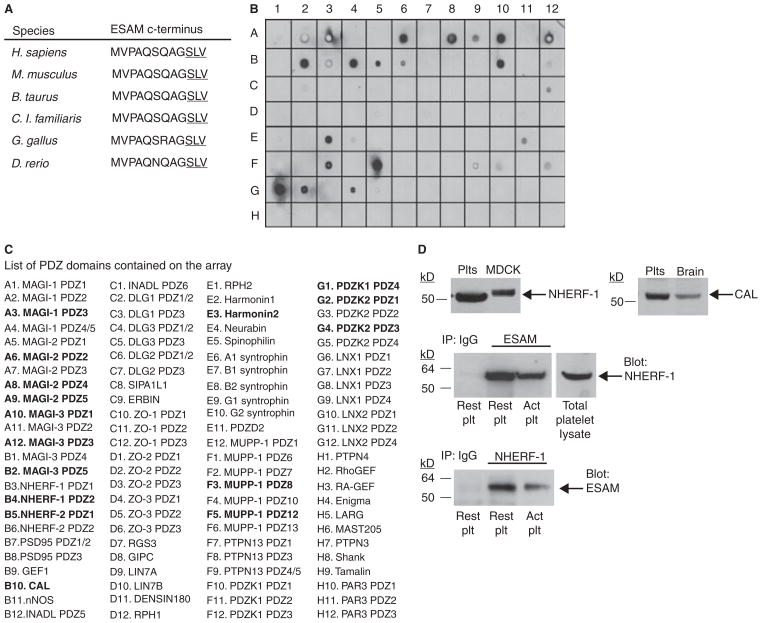
Methods and results: We found that following platelet activation ESAM localizes to the junctions between adjacent platelets, suggesting a role for this protein in contact-dependent events that regulate thrombus formation. To test this hypothesis we examined the effect of ESAM deletion on platelet function. In vivo, ESAM(-/-) mice achieved more stable hemostasis than wild-type mice following tail transection, and developed larger thrombi following laser injury of cremaster muscle arterioles. In vitro, ESAM(-/-) platelets aggregated at lower concentrations of G protein-dependent agonists than wild-type platelets, and were more resistant to disaggregation. In contrast, agonist-induced calcium mobilization, alpha(IIb)beta(3) activation, alpha-granule secretion and platelet spreading, were normal in ESAM-deficient platelets. To understand the molecular mechanism by which ESAM regulates platelet activity, we utilized a PDZ domain array to identify the scaffold protein NHERF-1 as an ESAM binding protein, and further demonstrated that it associates with ESAM in both resting and activated platelets.
Conclusions: These findings support a model in which ESAM localizes to platelet contacts following platelet activation in order to limit thrombus growth and stability so that the optimal hemostatic response occurs following vascular injury.
Conflict of interest statement
The authors state that they have no conflict of interest.
Figures

References
-
- Law DA, DeGuzman FR, Heiser P, Ministri-Madrid K, Killeen N, Phillips DR. Integrin cytoplasmic tyrosine motif is required for outside-in alphaIIbbeta3 signalling and platelet function. Nature. 1999;401:808–11. - PubMed
-
- Cosemans JM, Munnix IC, Wetzker R, Heller R, Jackson SP, Heemskerk JW. Continuous signaling via PI3K isoforms beta and gamma is required for platelet ADP receptor function in dynamic thrombus stabilization. Blood. 2006;108:3045–52. - PubMed
-
- Goto S, Tamura N, Ishida H, Ruggeri ZM. Dependence of platelet thrombus stability on sustained glycoprotein IIb/IIIa activation through adenosine 5′-diphosphate receptor stimulation and cyclic calcium signaling. J Am Coll Cardiol. 2006;47:155–62. - PubMed
-
- Andre P, Prasad KS, Denis CV, He M, Papalia JM, Hynes RO, Phillips DR, Wagner DD. CD40L stabilizes arterial thrombi by a beta3 integrin–dependent mechanism. Nat Med. 2002;8:247–52. - PubMed
-
- Angelillo-Scherrer A, Burnier L, Flores N, Savi P, DeMol M, Schaeffer P, Herbert JM, Lemke G, Goff SP, Matsushima GK, Earp HS, Vesin C, Hoylaerts MF, Plaisance S, Collen D, Conway EM, Wehrle-Haller B, Carmeliet P. Role of Gas6 receptors in platelet signaling during thrombus stabilization and implications for antithrombotic therapy. J Clin Invest. 2005;115:237–46. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
