

doi: 10.1128/JVI.01508-09.
Epub 2009 Sep 9.
Role of RNA structures in genome terminal sequences of the hepatitis C virus for replication and assembly
Affiliations
- PMID: 19740989
- PMCID: PMC2772684
- DOI: 10.1128/JVI.01508-09
Item in Clipboard
Role of RNA structures in genome terminal sequences of the hepatitis C virus for replication and assembly
J Virol.
2009 Nov.
Abstract
Hepatitis C virus (HCV) is a positive-strand RNA virus replicating its genome via a negative-strand [(-)] intermediate. Little is known about replication signals residing in the 3' end of HCV (-) RNA. Recent studies identified seven stem-loop structures (SL-I', -IIz', -IIy', -IIIa', -IIIb', -IIIcdef', and -IV') in this region. In the present study, we mapped the minimal region required for RNA replication to SL-I' and -IIz', functionally confirmed the SL-IIz' structure, and identified SL-IIIa' to -IV' as auxiliary replication elements. In addition, we show that the 5' nontranslated region of the genome most likely does not contain cis-acting RNA structures required for RNA packaging into infectious virions.
Figures
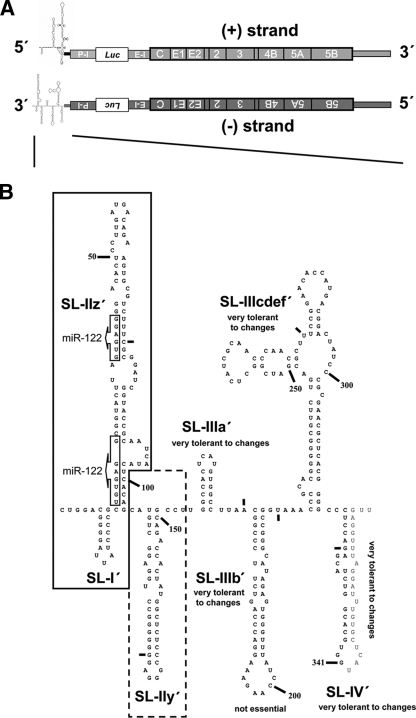
(A) Schematic presentation of the JFH1-derived reporter virus genome used throughout this study in the (+) and (−) orientations. Secondary structures of 5′ end of (+) RNA and 3′ end of (−) RNA are indicated. The poliovirus IRES (P-I) is positioned upstream of the firefly luciferase (Luc) reporter gene, and it is separated from the HCV 5′ NTR by a 61-nt-long spacer element (not indicated). Translation of the HCV coding region is mediated by the EMCV IRES (E-I). (B) Predicted RNA secondary structure of the 3′ end of JFH1 (−) RNA according to reference . The nomenclature of SL structures is given. Nucleotides are counted from the 3′ to 5′ direction; small bars indicate 25-nt intervals. Nucleotides downstream of position 341 are absent in the construct (see the supplemental material) and given only to highlight SL-IV′. Box arrows indicate complementary sequences of seed sequences for miRNA-122 binding in the 5′ NTR of (+) RNA. Minimal sequence required for RNA replication is framed; sequences that significantly contribute to RNA replication are framed with a dashed box.

Mutations affecting SL-I′ and SL-II′ in the 3′ end of (−) RNA and their impact on HCV RNA replication. (A) Detailed presentation of the mutants targeting the three 3′-terminal elements of (−) RNA. The wt structures are boxed with black lines; corresponding mutants are displayed next to each wt structure. Nucleotide substitutions are indicated in red, and their positions are highlighted with red lines; changes in the predicted RNA structure are indicated by dislocated sequences. The name of the mutant is given above each depicted RNA structure. Complementary sequences corresponding to seed sequences of miRNA-122 binding sites in the 5′ NTR of (+) RNA are boxed. Replication competence is summarized below each mutant. (B) Transient replication assays of mutants specified below each bar. In vitro transcripts were transfected into Huh7-Lunet cells (15) that were harvested 4, 24, 48, and 72 h posttransfection. RNA replication was scored by luciferase assay after normalization to the 4-h value reflecting transfection efficiency (15). Shown is the normalized 48-h luciferase value as a percentage of the normalized wt activity. We have shown earlier that the 48-h value reflects RNA replication competence best (6). GND, inactive replicon with a single amino acid substitution destroying the active site of the NS5B RNA polymerase. Mean values from three independent experiments are shown.

Mutations in SL-III′ and SL-IV′ and their impact on HCV RNA replication. (A) Detailed overview of constructed mutations. Nucleotide substitutions are highlighted with red letters, and their positions are emphasized with red lines; deletions are highlighted with blue letters and blue lines. Mutations predicted to disrupt proposed RNA elements are indicated by dislocation of the altered sequence. The wt structures are boxed with black lines; mutations affecting the same RNA element are given next to the corresponding wt structure. Names of mutants are given at the top of each panel, and replication competence is summarized below each mutant. (B) Replication competence of mutants that are specified below each bar. Mean values of three independent experiments are shown. For further details, see the legend to Fig. 2.

Mutations in the 5′ NTR of (+) RNA and their impact on production of infectious HCV particles. (A) Secondary structure of the 5′ NTR of JFH1 (+) wt RNA is framed (9). Nucleotide positions are indicated by small bars in 25-nt intervals; the start codon is circled. Nucleotides in blue indicate deletions, and their positions are further highlighted by blue lines; nucleotide substitutions are highlighted in red and with red lines. Possible rearrangements of RNA secondary structures induced by mutations are not considered. (B) Results of transient RNA replication assays are shown in the upper graph. Experimental conditions were the same as those described in the legend to Fig. 2, with the exception that Huh7.5 cells were used. Infectivity assays of the various mutants are shown in the lower panel. Forty-eight hours posttransfection, culture supernatants were harvested and used to infect naïve Huh7.5 cells. These were harvested 72 h after inoculation, and luciferase activities in cell lysates were determined. Values are normalized to the wt, which was set to 100%. Names of the constructs are displayed between both panels. Mutant nt 296 contains only the first 296 nt of the 5′ NTR and served as a reference; mutant Del E1/E2 is derived from the bicistronic wt genome, but lacks the envelope glycoproteins (27). This RNA replicates to wt levels, but does not release infectious particles and therefore was used to define background of the infectivity assay. Mean values of three independent experiments are shown.
References
-
- Bartenschlager, R., M. Frese, and T. Pietschmann. 2004. Novel insights into hepatitis C virus replication and persistence. Adv. Virus Res. 63:71-180. - PubMed
-
- Dutkiewicz, M., A. Swiatkowska, M. Figlerowicz, and J. Ciesiolka. 2008. Structural domains of the 3′-terminal sequence of the hepatitis C virus replicative strand. Biochemistry 47:12197-12207. - PubMed
-
- Fan, Z., Q. R. Yang, J. S. Twu, and A. H. Sherker. 1999. Specific in vitro association between the hepatitis C viral genome and core protein. J. Med. Virol. 59:131-134. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
