

Loss of the apical V-ATPase a-subunit VHA-6 prevents acidification of the intestinal lumen during a rhythmic behavior in C. elegans
- PMID: 19741196
- PMCID: PMC2777397
- DOI: 10.1152/ajpcell.00284.2009
Loss of the apical V-ATPase a-subunit VHA-6 prevents acidification of the intestinal lumen during a rhythmic behavior in C. elegans
Abstract
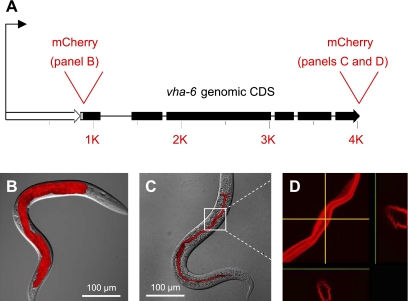
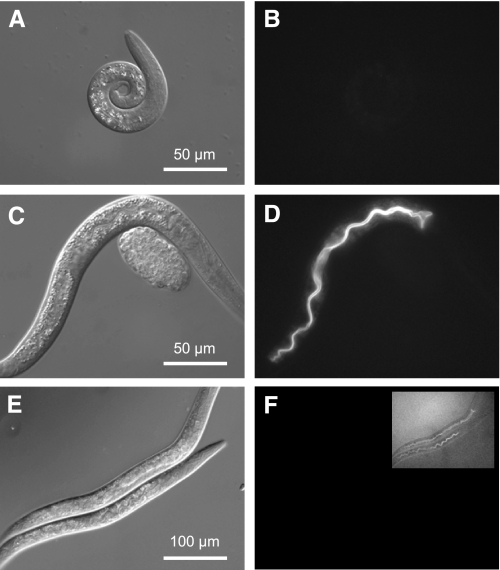
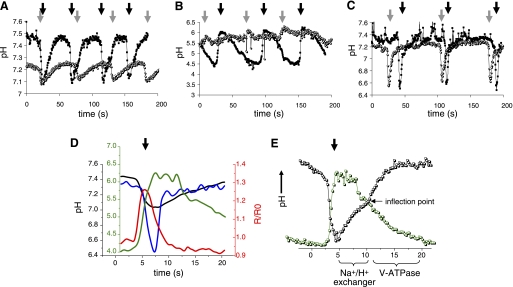
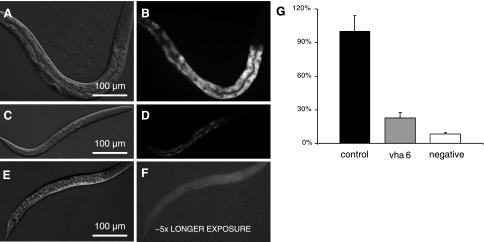
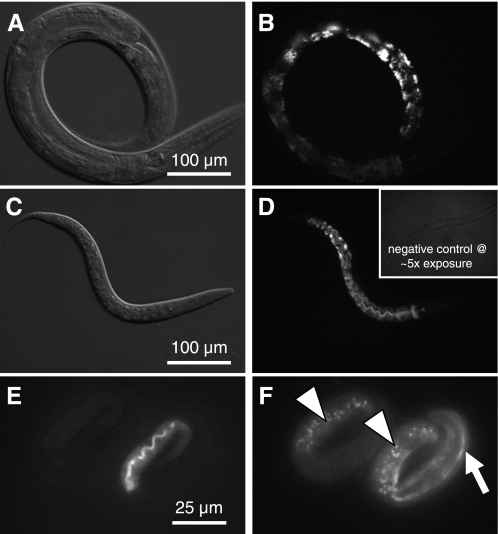
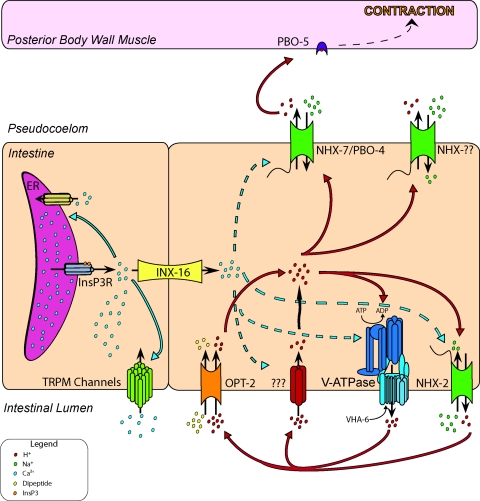
In Caenorhabditis elegans, oscillations of intestinal pH contribute to the rhythmic defecation behavior, but the acid-base transport mechanisms that facilitate proton movement are not well understood. Here, we demonstrate that VHA-6, an intestine-specific a-subunit of the H(+)-K(+)-ATPase complex (V-ATPase), resides in the apical membrane of the intestinal epithelial cells and is required for luminal acidification. Disruption of the vha-6 gene led to early developmental arrest; the arrest phenotype could be complemented by expression of a fluorescently labeled vha-6 transgene. To study the contribution of vha-6 to pH homeostasis in larval worms, we used a partial reduction of function through postembryonic single-generation RNA interference. We demonstrate that the inability to fully acidify the intestinal lumen coincides with a defect in pH recovery of the intestinal epithelial cells, suggesting that VHA-6 is essential for proton pumping following defecation. Moreover, intestinal dipeptide accumulation and fat storage are compromised by the loss of VHA-6, suggesting that luminal acidification promotes nutrient uptake in worms, as well as in mammals. Since acidified intracellular vesicles and autofluorescent storage granules are indistinguishable between the vha-6 mutant and controls, it is likely that the nutrient-restricted phenotype is due to a loss of plasma membrane V-ATPase activity specifically. These data establish a simple genetic model for proton pump-driven acidification. Since defecation occurs at 45-s intervals in worms, this model represents an opportunity to study acute regulation of V-ATPase activity on a short time scale and may be useful in the study of alternative treatments for acid-peptic disorders.
Figures

References
-
- Al-Awqati Q. Plasticity in epithelial polarity of renal intercalated cells: targeting of the H+-ATPase and band 3. Am J Physiol Cell Physiol 270: C1571–C1580, 1996 - PubMed
-
- Ashrafi K, Chang FY, Watts JL, Fraser AG, Kamath RS, Ahringer J, Ruvkun G. Genome-wide RNAi analysis of Caenorhabditis elegans fat regulatory genes. Nature 421: 268–272, 2003 - PubMed
-
- Beyenbach KW, Wieczorek H. The V-type H+ ATPase: molecular structure and function, physiological roles and regulation. J Exp Biol 209: 577–589, 2006 - PubMed
-
- Blair HC, Teitelbaum SL, Ghiselli R, Gluck S. Osteoclastic bone resorption by a polarized vacuolar proton pump. Science 245: 855–857, 1989 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases