

The G-quadruplex ligand telomestatin impairs binding of topoisomerase IIIalpha to G-quadruplex-forming oligonucleotides and uncaps telomeres in ALT cells
- PMID: 19742304
- PMCID: PMC2732903
- DOI: 10.1371/journal.pone.0006919
The G-quadruplex ligand telomestatin impairs binding of topoisomerase IIIalpha to G-quadruplex-forming oligonucleotides and uncaps telomeres in ALT cells
Abstract
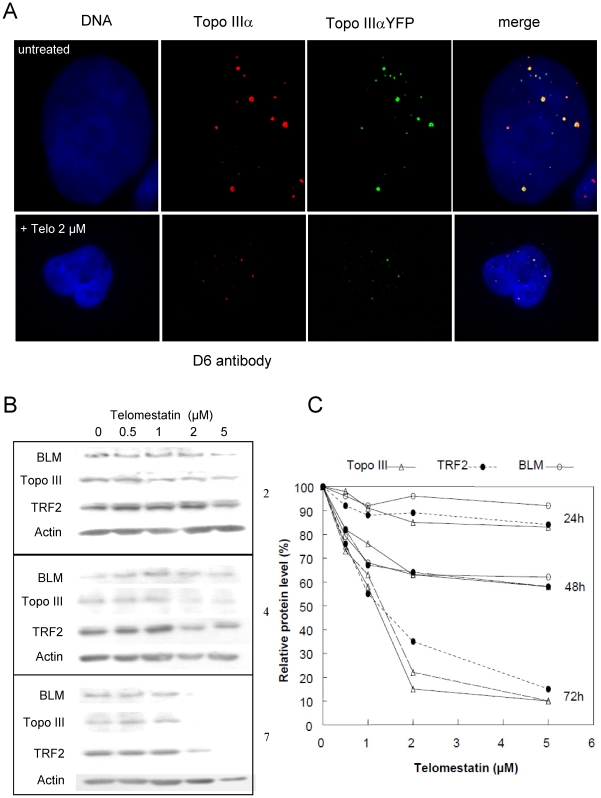
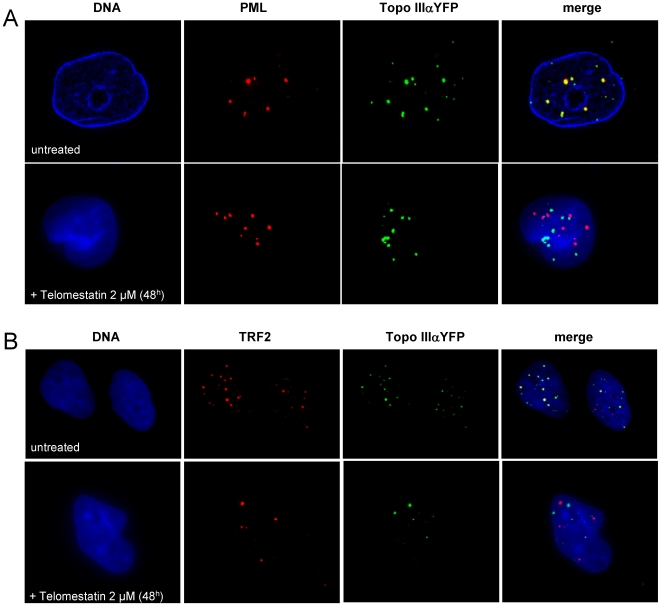
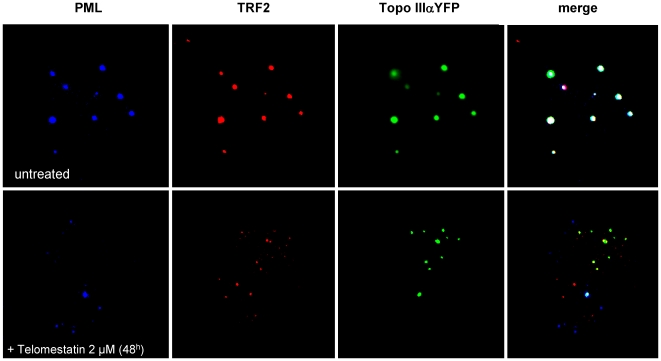
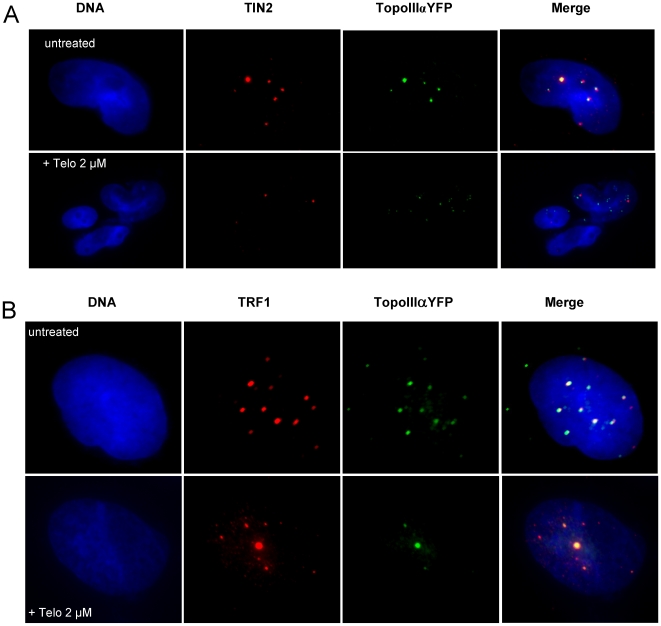
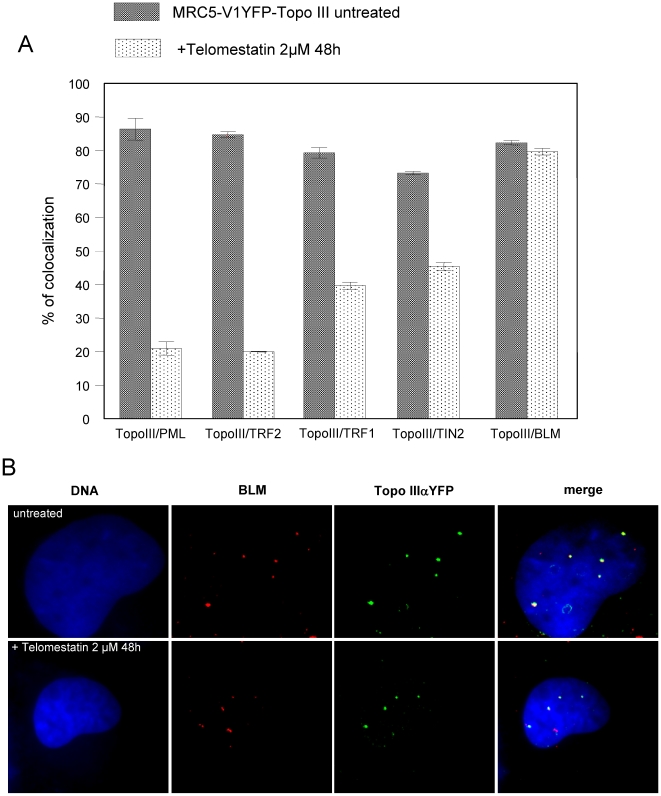
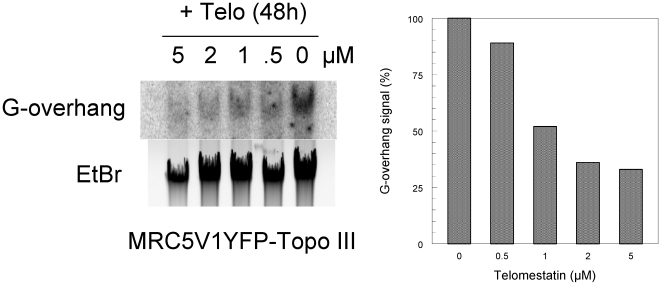
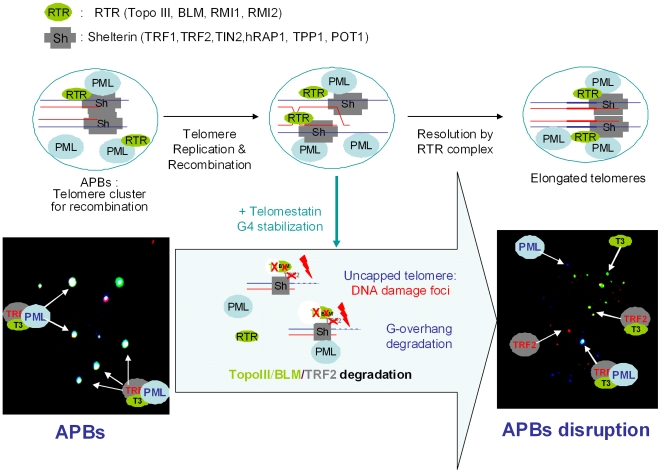
In Alternative Lengthening of Telomeres (ALT) cell lines, specific nuclear bodies called APBs (ALT-associated PML bodies) concentrate telomeric DNA, shelterin components and recombination factors associated with telomere recombination. Topoisomerase IIIalpha (Topo III) is an essential telomeric-associated factor in ALT cells. We show here that the binding of Topo III to telomeric G-overhang is modulated by G-quadruplex formation. Topo III binding to G-quadruplex-forming oligonucleotides was strongly inhibited by telomestatin, a potent and specific G-quadruplex ligand. In ALT cells, telomestatin treatment resulted in the depletion of the Topo III/BLM/TRF2 complex and the disruption of APBs and led to the segregation of PML, shelterin components and Topo III. Interestingly, a DNA damage response was observed at telomeres in telomestatin-treated cells. These data indicate the importance of G-quadruplex stabilization during telomere maintenance in ALT cells. The function of TRF2/Topo III/BLM in the resolution of replication intermediates at telomeres is discussed.
Conflict of interest statement
Figures

Similar articles
-
Topoisomerase IIIalpha is required for normal proliferation and telomere stability in alternative lengthening of telomeres.EMBO J. 2008 May 21;27(10):1513-24. doi: 10.1038/emboj.2008.74. Epub 2008 Apr 17. EMBO J. 2008. PMID: 18418389 Free PMC article.
-
The G-quadruplex ligand telomestatin inhibits POT1 binding to telomeric sequences in vitro and induces GFP-POT1 dissociation from telomeres in human cells.Cancer Res. 2006 Jul 15;66(14):6908-12. doi: 10.1158/0008-5472.CAN-06-1581. Cancer Res. 2006. PMID: 16849533
-
Telomestatin and diseleno sapphyrin bind selectively to two different forms of the human telomeric G-quadruplex structure.J Am Chem Soc. 2005 Jul 6;127(26):9439-47. doi: 10.1021/ja0505088. J Am Chem Soc. 2005. PMID: 15984871
-
Alternative lengthening of telomeres in mammalian cells.Oncogene. 2002 Jan 21;21(4):598-610. doi: 10.1038/sj.onc.1205058. Oncogene. 2002. PMID: 11850785 Review.
-
Telomere G-Quadruplex as a Potential Target to Accelerate Telomere Shortening by Expanding the Incomplete End-Replication of Telomere DNA.Curr Top Med Chem. 2015;15(19):1940-6. doi: 10.2174/1568026615666150515145552. Curr Top Med Chem. 2015. PMID: 25980416 Review.
Cited by
-
DNA folds threaten genetic stability and can be leveraged for chemotherapy.RSC Chem Biol. 2020 Sep 30;2(1):47-76. doi: 10.1039/d0cb00151a. eCollection 2021 Feb 1. RSC Chem Biol. 2020. PMID: 35340894 Free PMC article. Review.
-
Effects of Replication and Transcription on DNA Structure-Related Genetic Instability.Genes (Basel). 2017 Jan 5;8(1):17. doi: 10.3390/genes8010017. Genes (Basel). 2017. PMID: 28067787 Free PMC article. Review.
-
Fundamentals of G-quadruplex biology.Annu Rep Med Chem. 2020;54:3-44. doi: 10.1016/bs.armc.2020.06.004. Epub 2020 Jul 30. Annu Rep Med Chem. 2020. PMID: 32836507 Free PMC article.
-
G-quadruplex Stabilization Fuels the ALT Pathway in ALT-positive Osteosarcoma Cells.Genes (Basel). 2020 Mar 13;11(3):304. doi: 10.3390/genes11030304. Genes (Basel). 2020. PMID: 32183119 Free PMC article.
-
Cation involvement in telomestatin binding to g-quadruplex DNA.J Nucleic Acids. 2010;2010:121259. doi: 10.4061/2010/121259. Epub 2010 Jun 16. J Nucleic Acids. 2010. PMID: 20700418 Free PMC article.
References
-
- Palm W, de Lange T. How shelterin protects Mammalian telomeres. Annu Rev Genet. 2008;42:301–334. - PubMed
-
- McEachern MJ, Krauskopf A, Blackburn EH. Telomeres and their control. Annu Rev Genet. 2000;34:331–358. - PubMed
-
- Cesare AJ, Reddel RR. Telomere uncapping and alternative lengthening of telomeres. Mech Ageing Dev. 2008;129:99–108. - PubMed
-
- Huppert JL. Hunting G-quadruplexes. Biochimie. 2008;90:1140–1148. - PubMed
-
- Monchaud D, Teulade-Fichou MP. A hitchhiker's guide to G-quadruplex ligands. Org Biomol Chem. 2008;6:627–636. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
