

Limited expansion of virus-specific CD8 T cells in the aged environment
- PMID: 19744506
- PMCID: PMC2839881
- DOI: 10.1016/j.mad.2009.08.007
Limited expansion of virus-specific CD8 T cells in the aged environment
Abstract
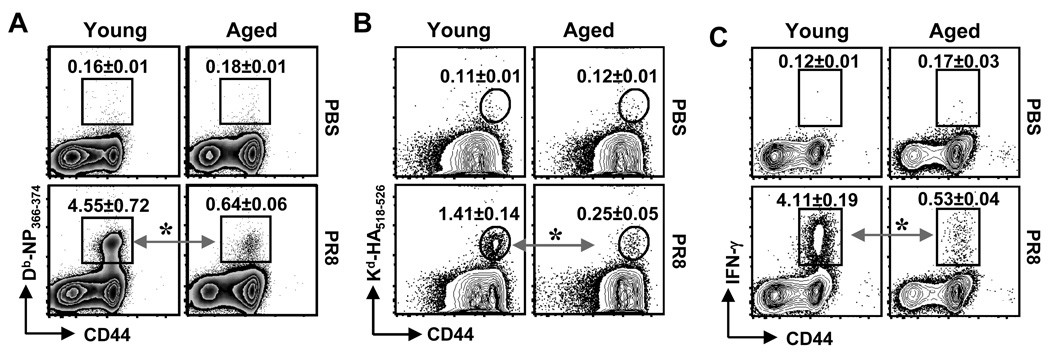
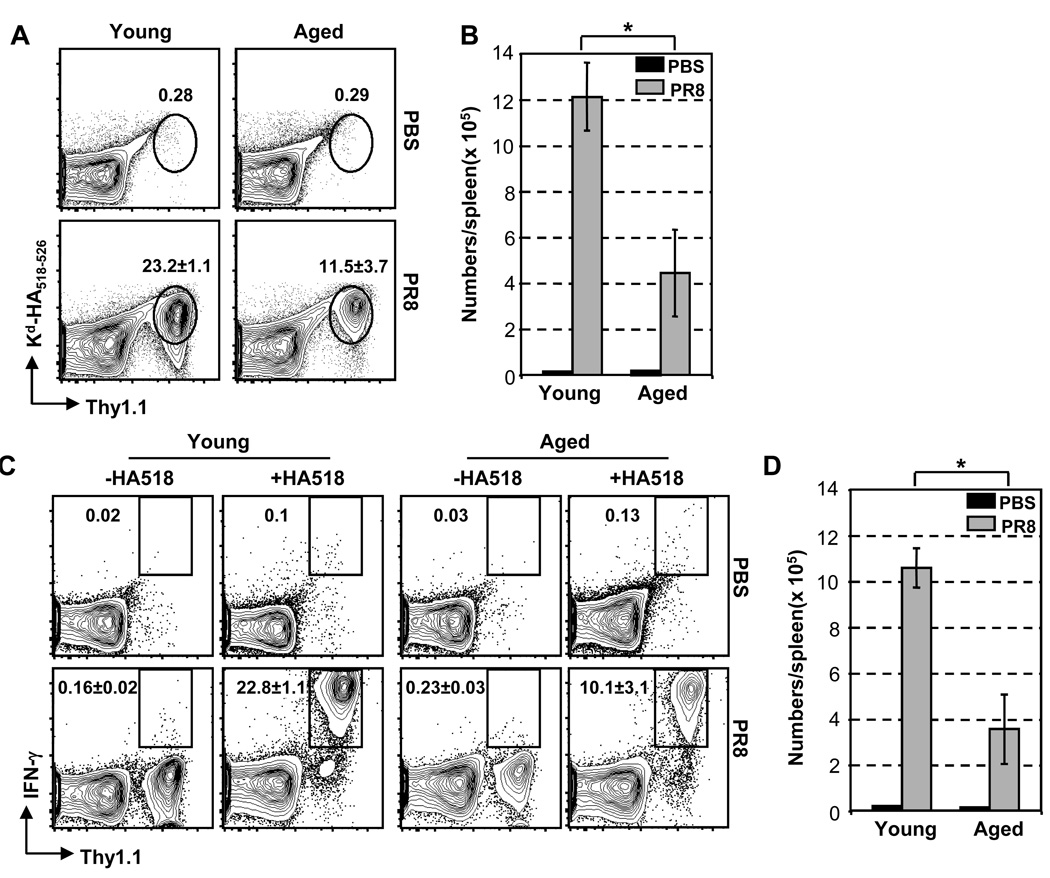
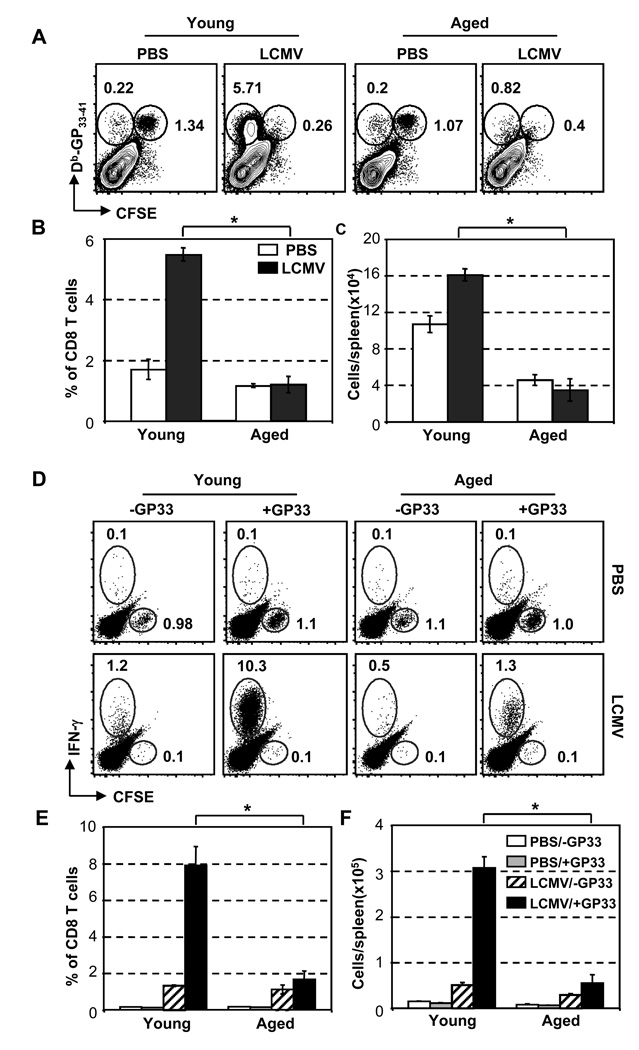
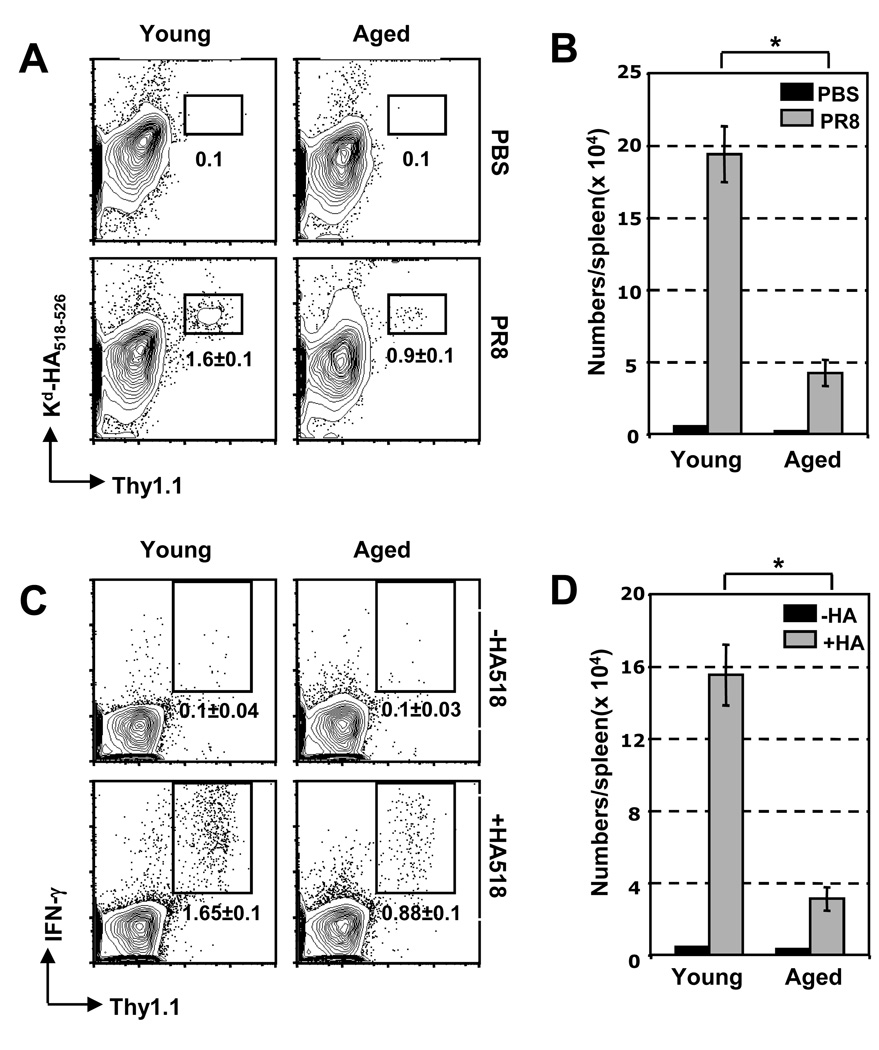
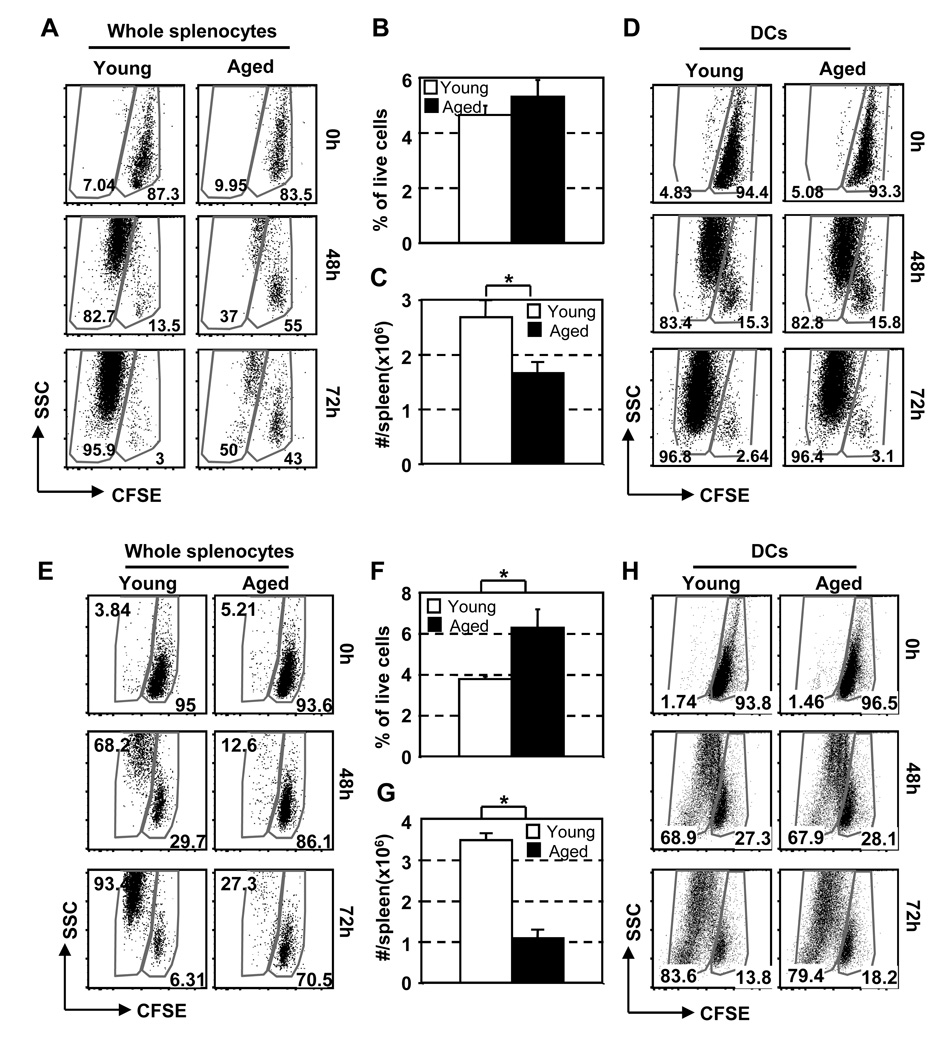
The mechanisms responsible for the diminished immune response seen with aging are unclear. In this study, we investigate the contributions of alterations in the lymphoid microenvironment to this decrease. Using adoptive transfer of virus-specific transgenic CD8 T cells, we demonstrate that the aged environment inhibits the clonal expansion of specific CD8 T cells from young mice during virus infection. Transferred specific CD8 T cells from young mice demonstrated a response reflecting the CD8 T cell response of the intact aged host: the CD8 T cells expand more slowly and have a decreased maximal expansion in an aged compared to a young environment. While isolated DCs (MHC II(+) CD11c(+)) of aged mice maintain their ability to support CD8 T cell Ag-specific expansion in vitro, splenocytes demonstrated an age-associated decrease in this ability. Since the percentages of various populations of DCs in splenocytes demonstrate no significant alteration with age, this diminished APC activity of splenocytes of aged mice may reflect inhibitory activity of other cell populations. The results of this study demonstrate that elements of the aged environment play an important role in the alteration of T cell response to virus infection in the aged.
Figures
References
-
- Appay V, Rowland-Jones SL. The assessment of antigen-specific CD8+ T cells through the combination of MHC class I tetramer and intracellular staining. J. Immunol. Methods. 2002;268:9–19. - PubMed
-
- Ashton-Rickardt PG, Bandeira A, Delaney JR, Van Kaer L, Pircher HP, Zinkernagel RM, Tonegawa S. Evidence for a differential avidity model of T cell selection in the thymus. Cell. 1994;76:651–663. - PubMed
-
- Beharka AA, Wu D, Han SN, Meydani SN. Macrophage prostaglandin production contributes to the age-associated decrease in T cell function which is reversed by the dietary antioxidant vitamin E. Mech. Ageing Dev. 1997;93:59–77. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
