

Human Naa50p (Nat5/San) displays both protein N alpha- and N epsilon-acetyltransferase activity
- PMID: 19744929
- PMCID: PMC2781511
- DOI: 10.1074/jbc.M109.001347
Human Naa50p (Nat5/San) displays both protein N alpha- and N epsilon-acetyltransferase activity
Abstract
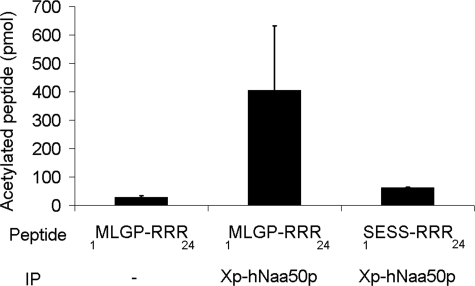
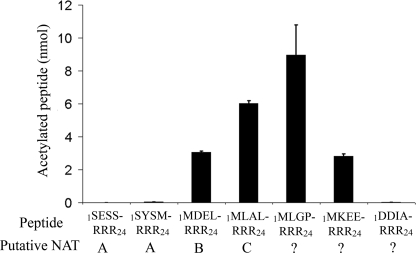
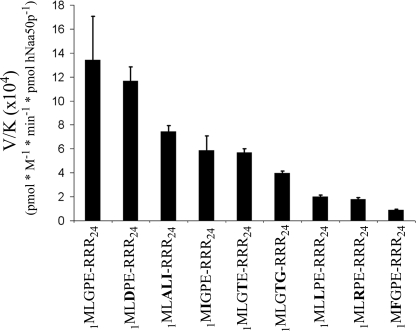
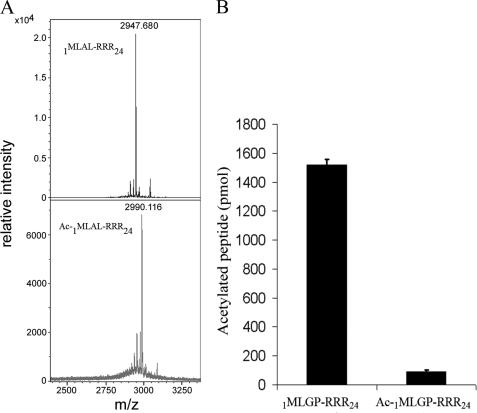
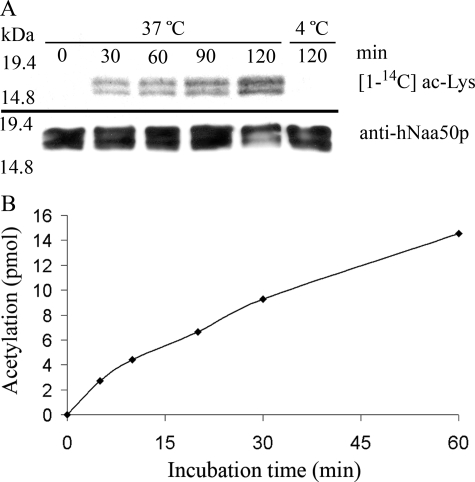
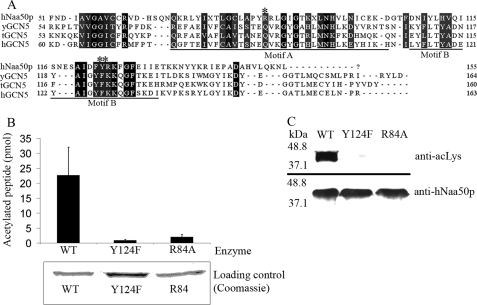
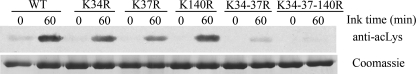
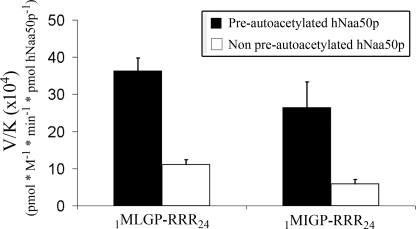
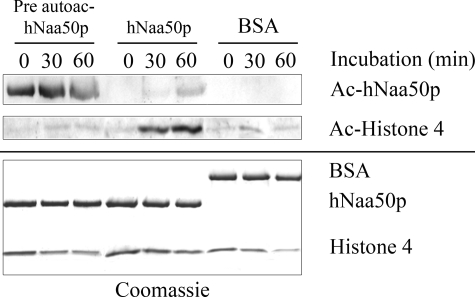
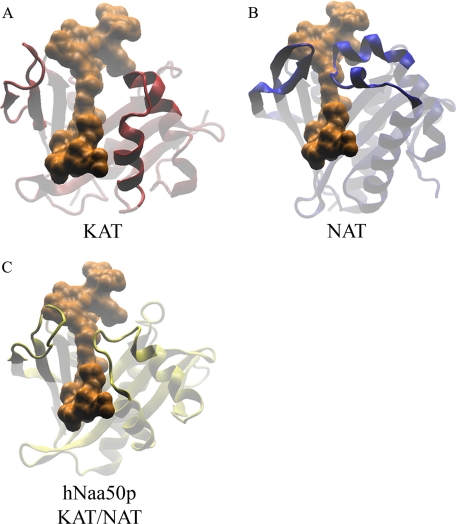
Protein acetylation is a widespread modification that is mediated by site-selective acetyltransferases. KATs (lysine N(epsilon)-acetyltransferases), modify the side chain of specific lysines on histones and other proteins, a central process in regulating gene expression. N(alpha)-terminal acetylation occurs on the ribosome where the alpha amino group of nascent polypeptides is acetylated by NATs (N-terminal acetyltransferase). In yeast, three different NAT complexes were identified NatA, NatB, and NatC. NatA is composed of two main subunits, the catalytic subunit Naa10p (Ard1p) and Naa15p (Nat1p). Naa50p (Nat5) is physically associated with NatA. In man, hNaa50p was shown to have acetyltransferase activity and to be important for chromosome segregation. In this study, we used purified recombinant hNaa50p and multiple oligopeptide substrates to identify and characterize an N(alpha)-acetyltransferase activity of hNaa50p. As the preferred substrate this activity acetylates oligopeptides with N termini Met-Leu-Xxx-Pro. Furthermore, hNaa50p autoacetylates lysines 34, 37, and 140 in vitro, modulating hNaa50p substrate specificity. In addition, histone 4 was detected as a hNaa50p KAT substrate in vitro. Our findings thus provide the first experimental evidence of an enzyme having both KAT and NAT activities.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
