

A dimeric structure for archaeal box C/D small ribonucleoproteins
- PMID: 19745151
- PMCID: PMC2975540
- DOI: 10.1126/science.1176099
A dimeric structure for archaeal box C/D small ribonucleoproteins
Abstract
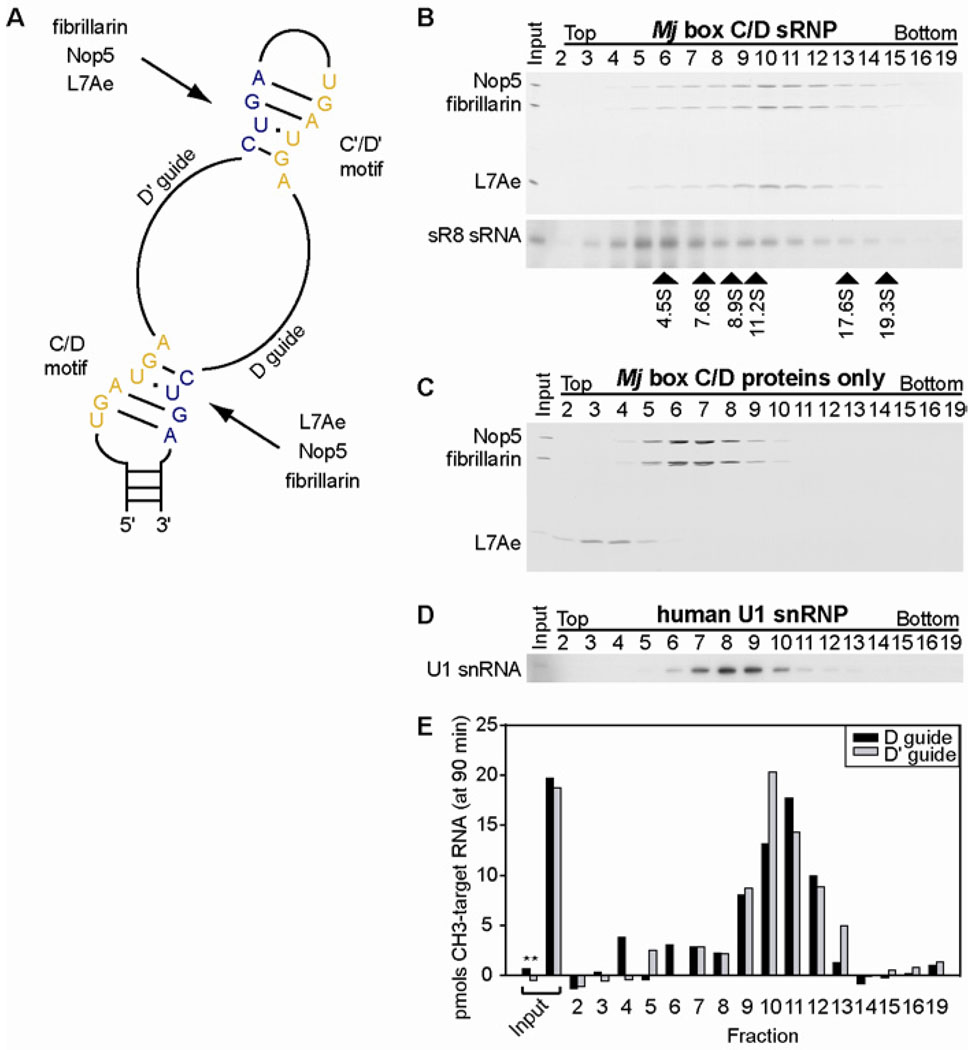
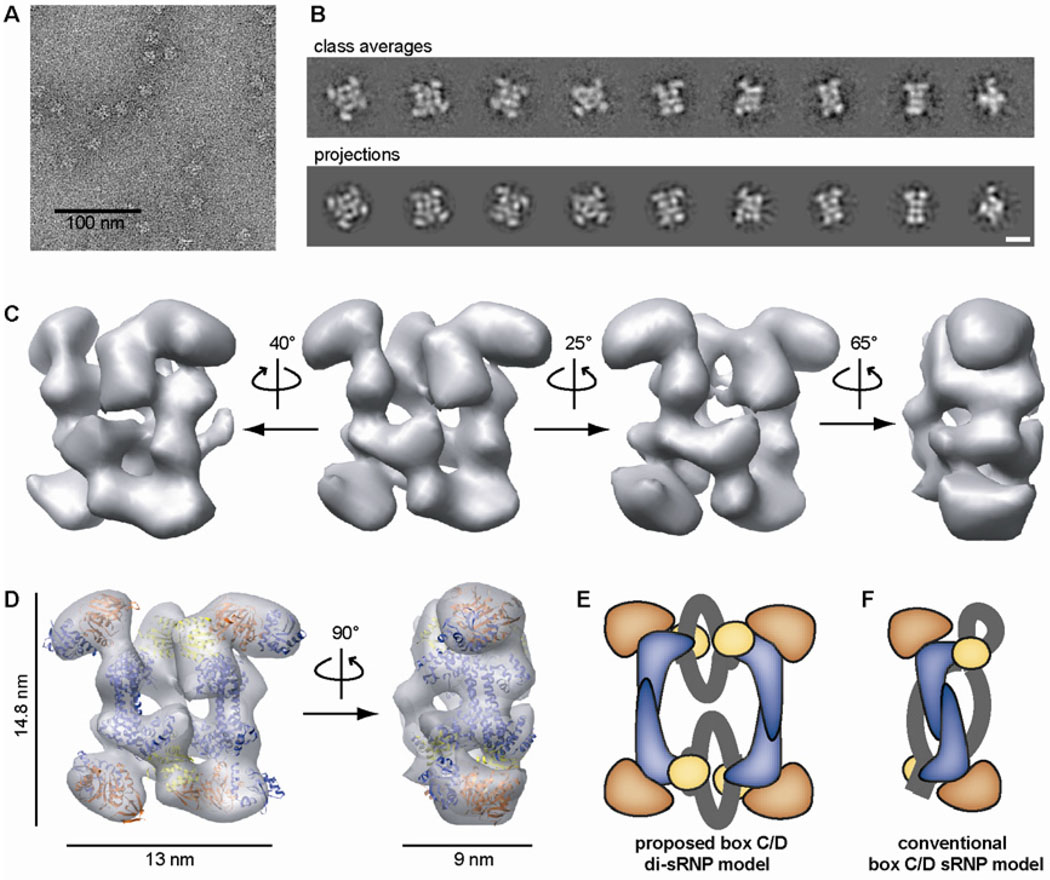
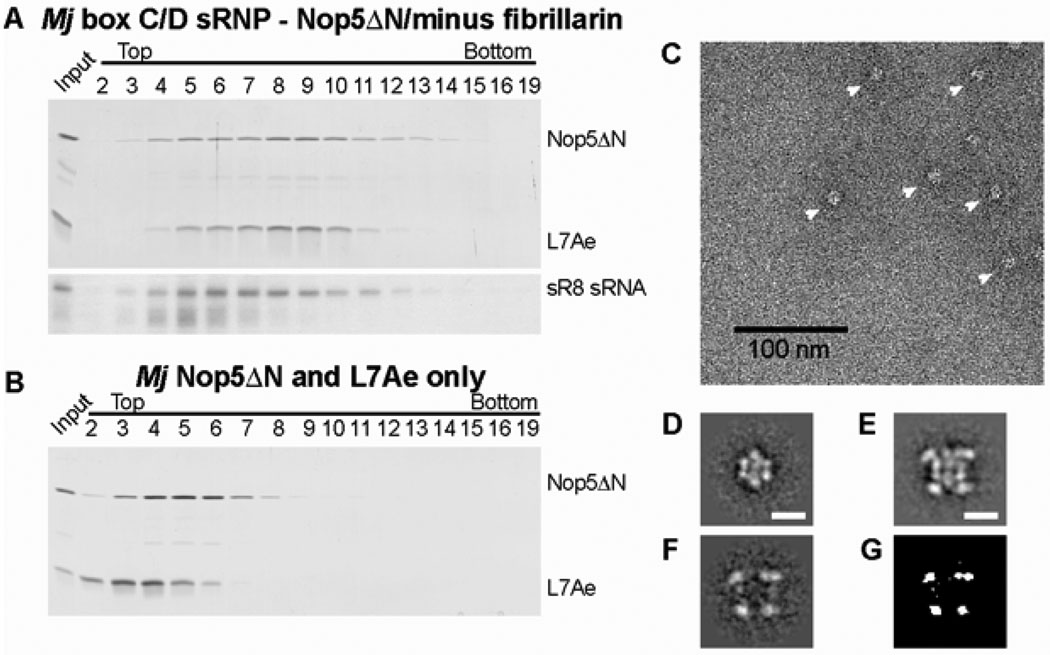
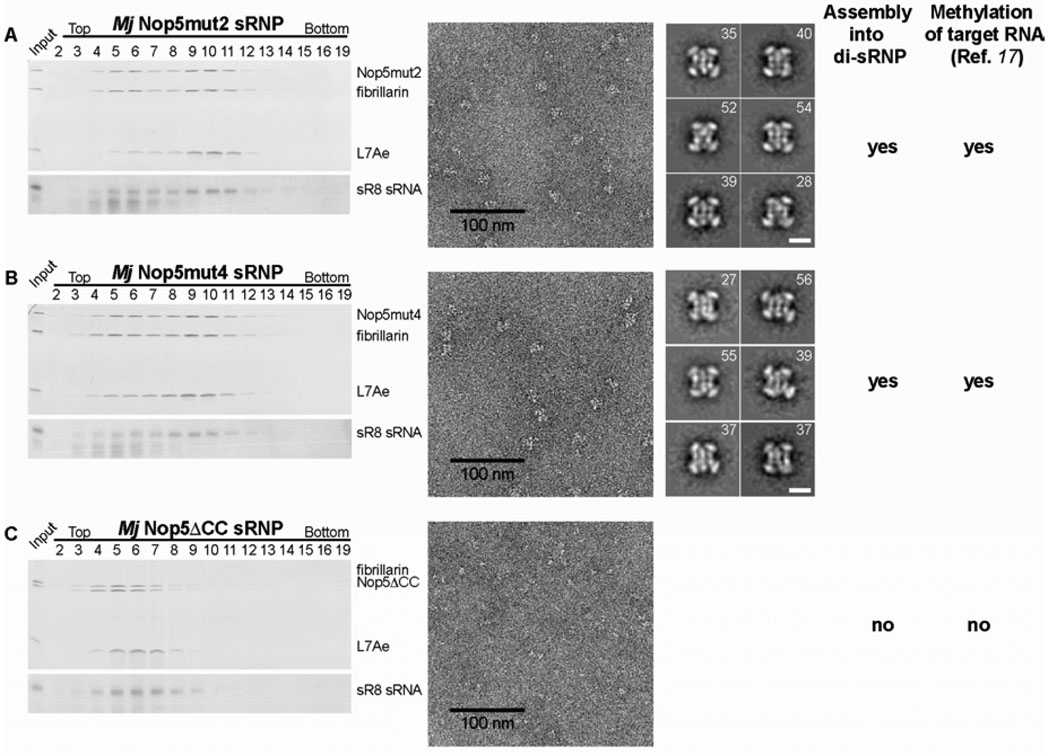
Methylation of ribosomal RNA (rRNA) is required for optimal protein synthesis. Multiple 2'-O-ribose methylations are carried out by box C/D guide ribonucleoproteins [small ribonucleoproteins (sRNPs) and small nucleolar ribonucleoproteins (snoRNPs)], which are conserved from archaea to eukaryotes. Methylation is dictated by base pairing between the specific guide RNA component of the sRNP or snoRNP and the target rRNA. We determined the structure of a reconstituted and catalytically active box C/D sRNP from the archaeon Methanocaldococcus jannaschii by single-particle electron microscopy. We found that archaeal box C/D sRNPs unexpectedly formed a dimeric structure with an alternative organization of their RNA and protein components that challenges the conventional view of their architecture. Mutational analysis demonstrated that this di-sRNP structure was relevant for the enzymatic function of archaeal box C/D sRNPs.
Figures
References
-
- Decatur WA, Fournier MJ. Trends Biochem Sci. 2002;27:344. - PubMed
-
- Liang XH, Liu Q, Fournier MJ. Mol Cell. 2007;28:965. - PubMed
-
- Kiss-Laszlo Z, Henry Y, Bachellerie JP, Caizergues-Ferrer M, Kiss T. Cell. 1996;85:1077. - PubMed
-
- Omer AD, et al. Science. 2000;288:517. - PubMed
-
- Matera AG, Terns RM, Terns MP. Nat Rev Mol Cell Biol. 2007;8:209. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
