

Decreased expression and activity of cAMP phosphodiesterases in cardiac hypertrophy and its impact on beta-adrenergic cAMP signals
- PMID: 19745166
- PMCID: PMC2792993
- DOI: 10.1161/CIRCRESAHA.109.197947
Decreased expression and activity of cAMP phosphodiesterases in cardiac hypertrophy and its impact on beta-adrenergic cAMP signals
Abstract
Rationale: Multiple cyclic nucleotide phosphodiesterases (PDEs) degrade cAMP in cardiomyocytes but the role of PDEs in controlling cAMP signaling during pathological cardiac hypertrophy is poorly defined.
Objective: Evaluate the beta-adrenergic regulation of cardiac contractility and characterize the changes in cardiomyocyte cAMP signals and cAMP-PDE expression and activity following cardiac hypertrophy.
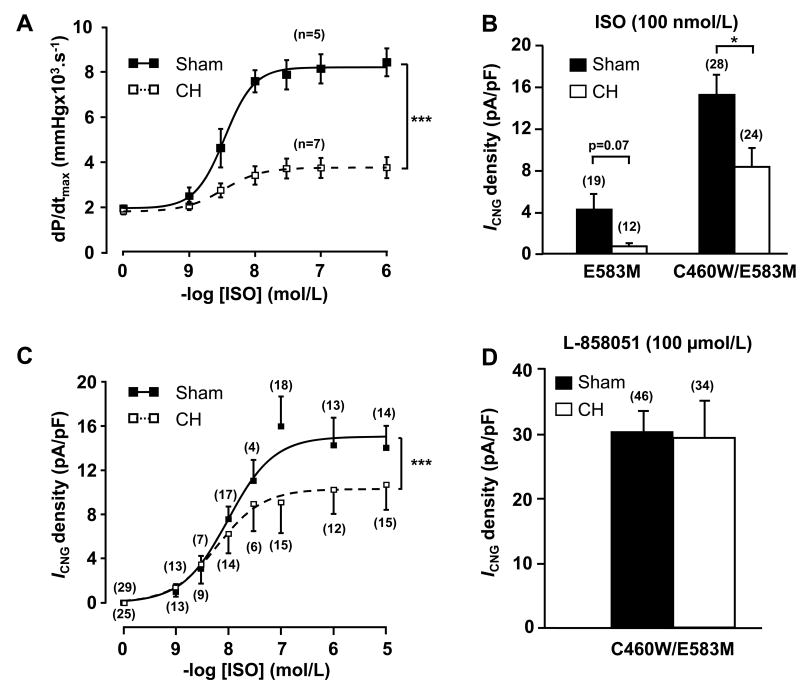
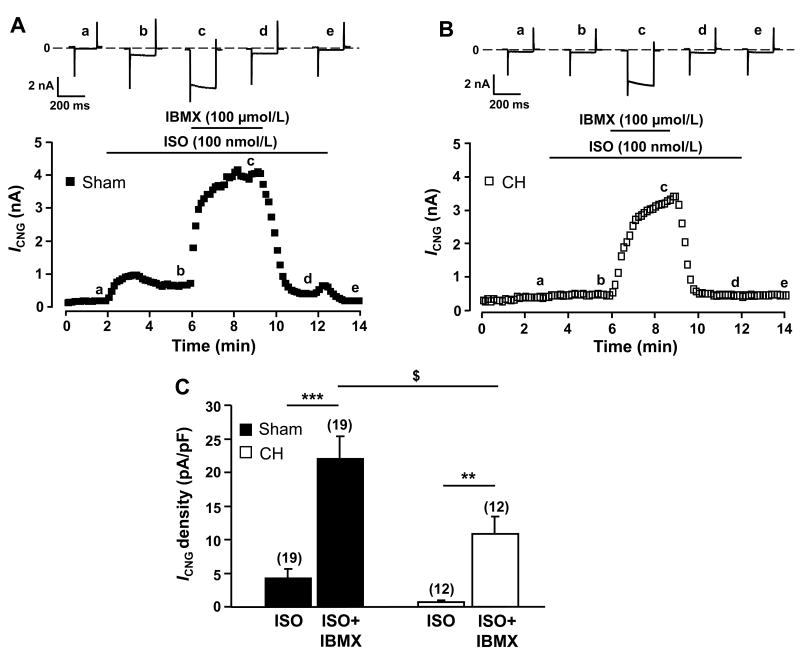
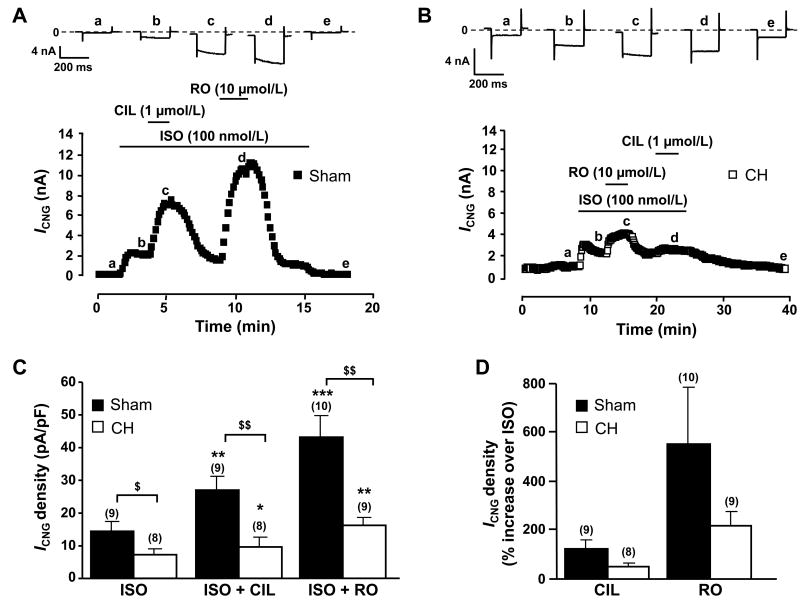
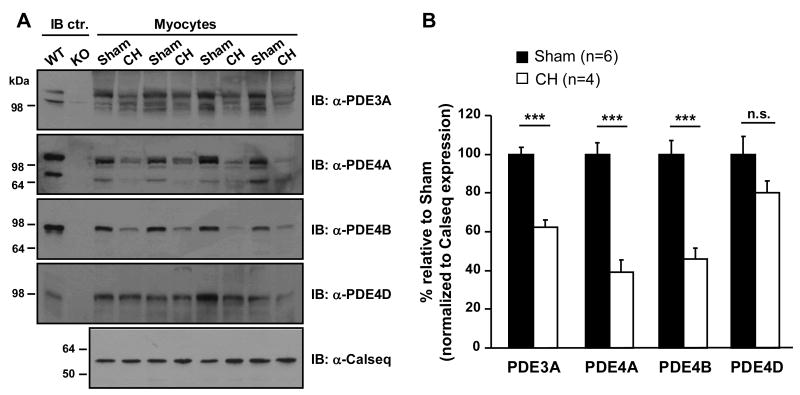
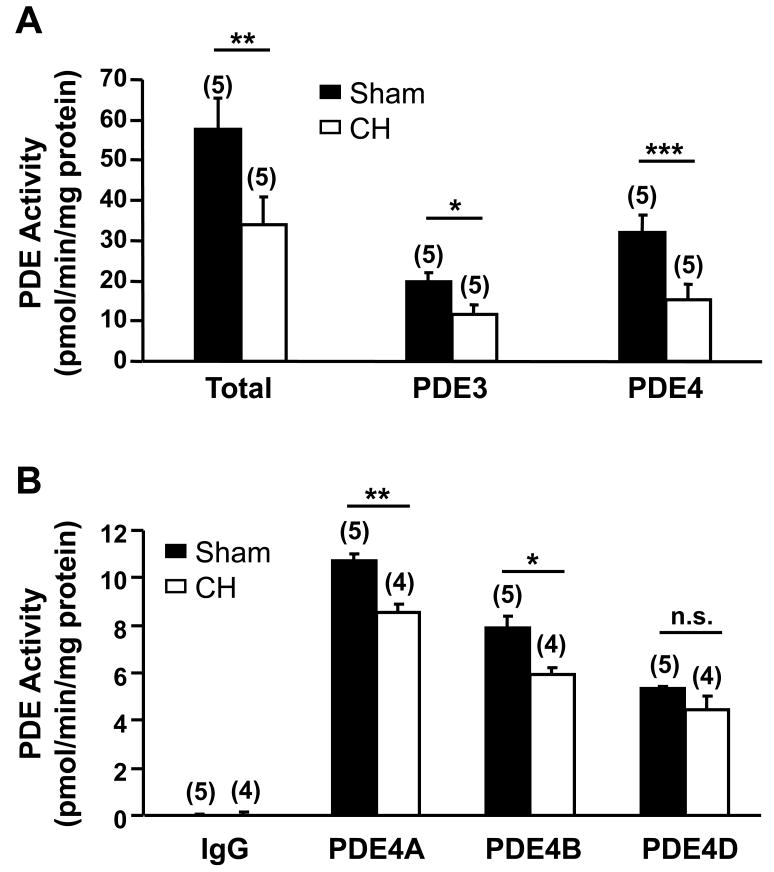
Methods and results: Cardiac hypertrophy was induced in rats by thoracic aortic banding over a time period of 5 weeks and was confirmed by anatomic measurements and echocardiography. Ex vivo myocardial function was evaluated in Langendorff-perfused hearts. Engineered cyclic nucleotide-gated (CNG) channels were expressed in single cardiomyocytes to monitor subsarcolemmal cAMP using whole-cell patch-clamp recordings of the associated CNG current (I(CNG)). PDE variant activity and protein level were determined in purified cardiomyocytes. Aortic stenosis rats exhibited a 67% increase in heart weight compared to sham-operated animals. The inotropic response to maximal beta-adrenergic stimulation was reduced by approximately 54% in isolated hypertrophied hearts, along with a approximately 32% decrease in subsarcolemmal cAMP levels in hypertrophied myocytes. Total cAMP hydrolytic activity as well as PDE3 and PDE4 activities were reduced in hypertrophied myocytes, because of a reduction of PDE3A, PDE4A, and PDE4B, whereas PDE4D was unchanged. Regulation of beta-adrenergic cAMP signals by PDEs was blunted in hypertrophied myocytes, as demonstrated by the diminished effects of IBMX (100 micromol/L) and of both the PDE3 inhibitor cilostamide (1 micromol/L) and the PDE4 inhibitor Ro 201724 (10 micromol/L).
Conclusions: Beta-adrenergic desensitization is accompanied by a reduction in cAMP-PDE and an altered modulation of beta-adrenergic cAMP signals in cardiac hypertrophy.
Conflict of interest statement
All authors declare no conflict of interest.
Figures
References
-
- Bers DM. Cardiac excitation-contraction coupling. Nature. 2002;415:198–205. - PubMed
-
- Lugnier C. Cyclic nucleotide phosphodiesterase (PDE) superfamily: a new target for the development of specific therapeutic agents. Pharmacol Ther. 2006;109:366–398. - PubMed
-
- Fischmeister R, Castro LRV, Abi-Gerges A, Rochais F, Jurevicˇius J, Leroy J, Vandecasteele G. Compartmentation of cyclic nucleotide signaling in the heart: The role of cyclic nucleotide phosphodiesterases. Circ Res. 2006;99:816–828. - PubMed
-
- Mongillo M, McSorley T, Evellin S, Sood A, Lissandron V, Terrin A, Huston E, Hannawacker A, Lohse MJ, Pozzan T, Houslay MD, Zaccolo M. Fluorescence resonance energy transfer-based analysis of cAMP dynamics in live neonatal rat cardiac myocytes reveals distinct functions of compartmentalized phosphodiesterases. Circ Res. 2004;95:65–75. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
