

Pre-fibrillar alpha-synuclein variants with impaired beta-structure increase neurotoxicity in Parkinson's disease models
- PMID: 19745811
- PMCID: PMC2771093
- DOI: 10.1038/emboj.2009.257
Pre-fibrillar alpha-synuclein variants with impaired beta-structure increase neurotoxicity in Parkinson's disease models
Abstract
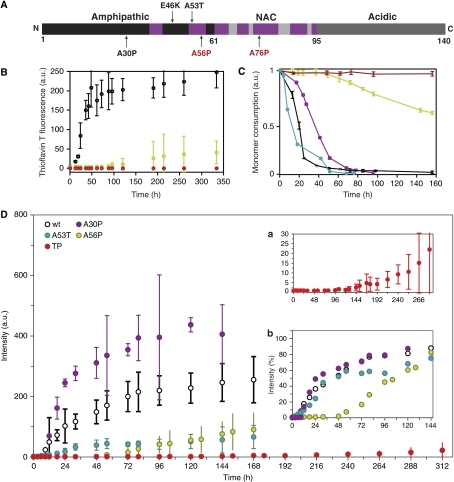
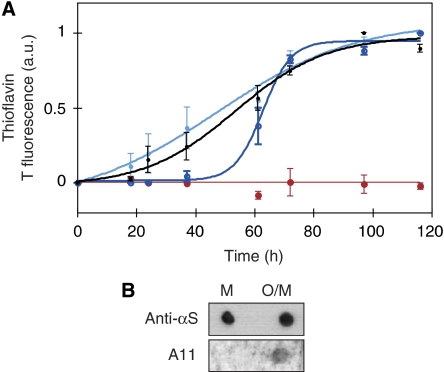
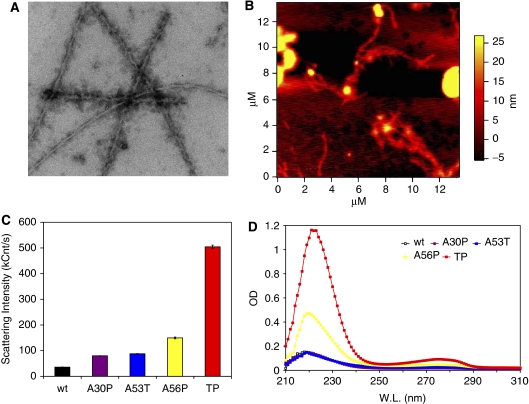
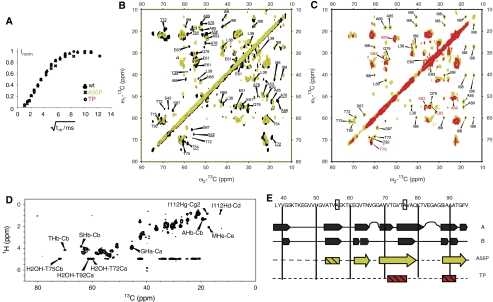
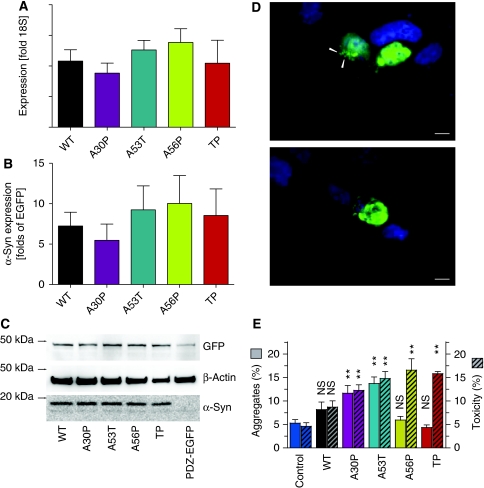
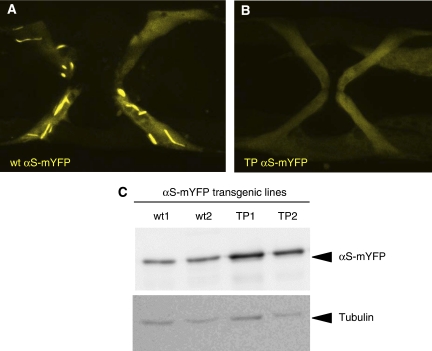
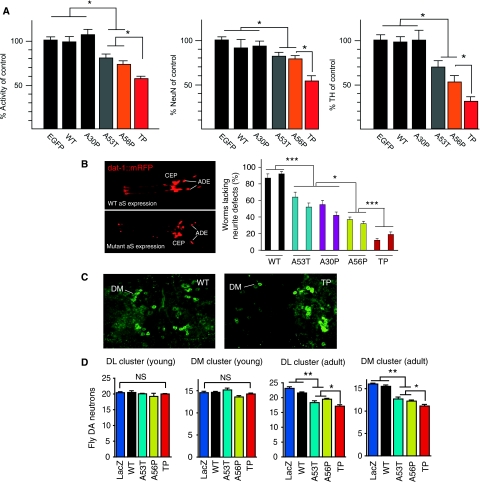
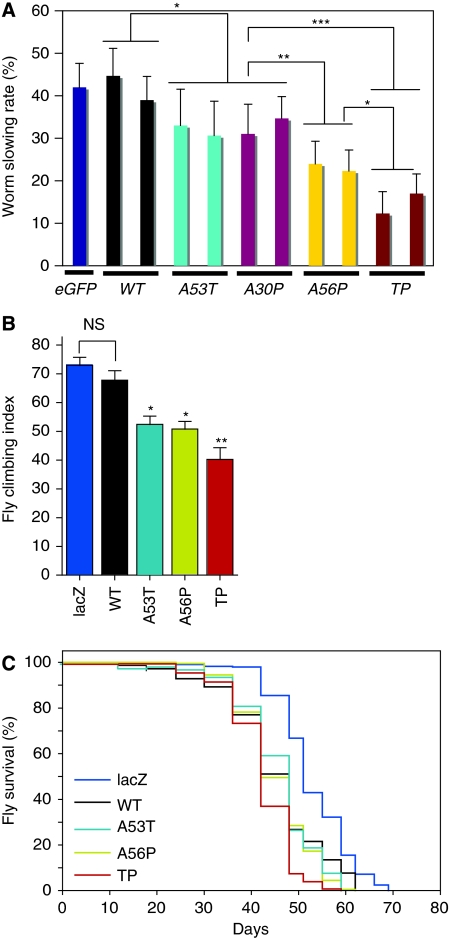
The relation of alpha-synuclein (alphaS) aggregation to Parkinson's disease (PD) has long been recognized, but the mechanism of toxicity, the pathogenic species and its molecular properties are yet to be identified. To obtain insight into the function different aggregated alphaS species have in neurotoxicity in vivo, we generated alphaS variants by a structure-based rational design. Biophysical analysis revealed that the alphaS mutants have a reduced fibrillization propensity, but form increased amounts of soluble oligomers. To assess their biological response in vivo, we studied the effects of the biophysically defined pre-fibrillar alphaS mutants after expression in tissue culture cells, in mammalian neurons and in PD model organisms, such as Caenorhabditis elegans and Drosophila melanogaster. The results show a striking correlation between alphaS aggregates with impaired beta-structure, neuronal toxicity and behavioural defects, and they establish a tight link between the biophysical properties of multimeric alphaS species and their in vivo function.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
-
- Ader C, Schneider R, Seidel K, Etzkorn M, Becker S, Baldus M (2009) Structural rearrangements of membrane proteins probed by water-edited solid-state NMR spectroscopy. J Am Chem Soc 131: 170–176 - PubMed
-
- Chen L, Feany MB (2005) Alpha-synuclein phosphorylation controls neurotoxicity and inclusion formation in a Drosophila model of Parkinson disease. Nat Neurosci 8: 657–663 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
