

Quinolones: action and resistance updated
- PMID: 19747119
- PMCID: PMC3182077
- DOI: 10.2174/156802609789630947
Quinolones: action and resistance updated
Abstract
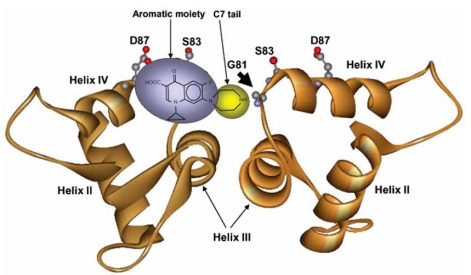
The quinolones trap DNA gyrase and DNA topoisomerase IV on DNA as complexes in which the DNA is broken but constrained by protein. Early studies suggested that drug binding occurs largely along helix-4 of the GyrA (gyrase) and ParC (topoisomerase IV) proteins. However, recent X-ray crystallography shows drug intercalating between the -1 and +1 nucleotides of cut DNA, with only one end of the drug extending to helix-4. These two models may reflect distinct structural steps in complex formation. A consequence of drug-enzyme-DNA complex formation is reversible inhibition of DNA replication; cell death arises from subsequent events in which bacterial chromosomes are fragmented through two poorly understood pathways. In one pathway, chromosome fragmentation stimulates excessive accumulation of highly toxic reactive oxygen species that are responsible for cell death. Quinolone resistance arises stepwise through selective amplification of mutants when drug concentrations are above the MIC and below the MPC, as observed with static agar plate assays, dynamic in vitro systems, and experimental infection of rabbits. The gap between MIC and MPC can be narrowed by compound design that should restrict the emergence of resistance. Resistance is likely to become increasingly important, since three types of plasmid-borne resistance have been reported.
Figures



References
-
- Kato J-I, Nishimura Y, Imamura R, Niki H, Hiraga S, Suzuki H. New topoisomerase essential for chromosome segregation in E. coli. Cell. 1990;63:393–404. - PubMed
-
- Hiasa H, DiGate R, Marians K. Decatenating activity of Escherichia coli DNA gyrase and topoisomerases I and III during oriC and pBR322 DNA replication in vitro. J. Biol. Chem. 1994;269:2093–2099. - PubMed
-
- Kreuzer KN, Cozzarelli NR. Formation and resolution of DNA catenanes by DNA gyrase. Cell. 1980;20:245–254. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
