

Mortality shifts in Caenorhabditis elegans: remembrance of conditions past
- PMID: 19747231
- PMCID: PMC2784026
- DOI: 10.1111/j.1474-9726.2009.00523.x
Mortality shifts in Caenorhabditis elegans: remembrance of conditions past
Abstract
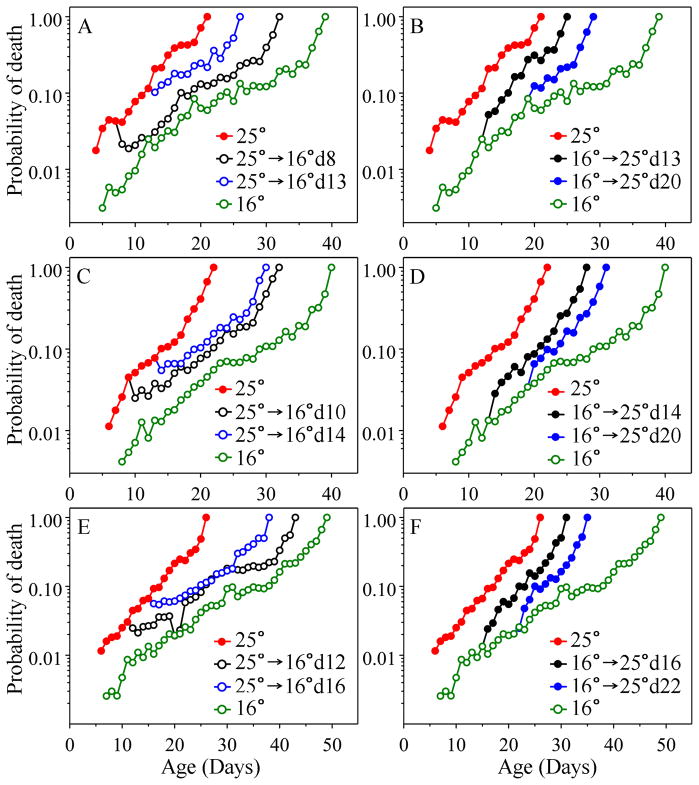
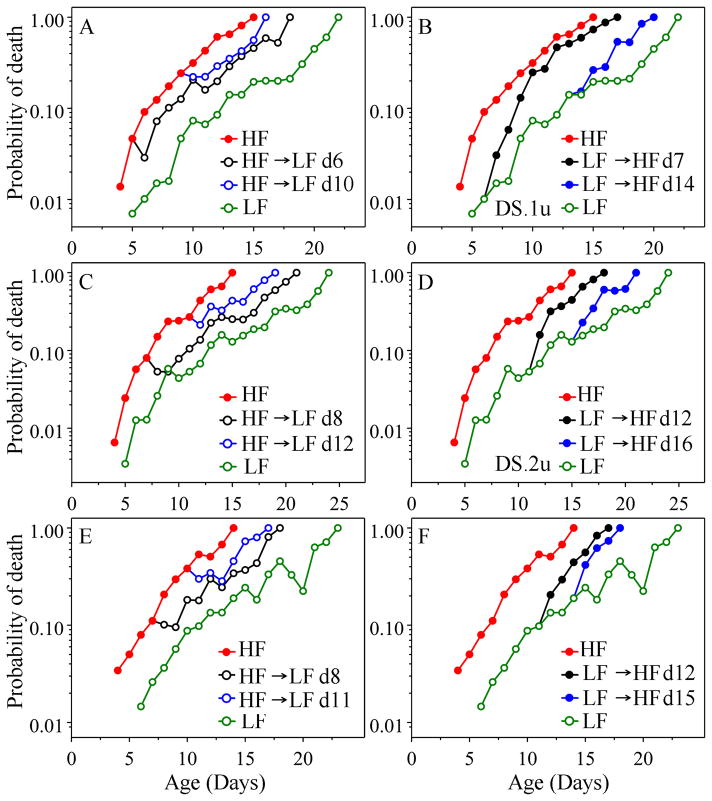
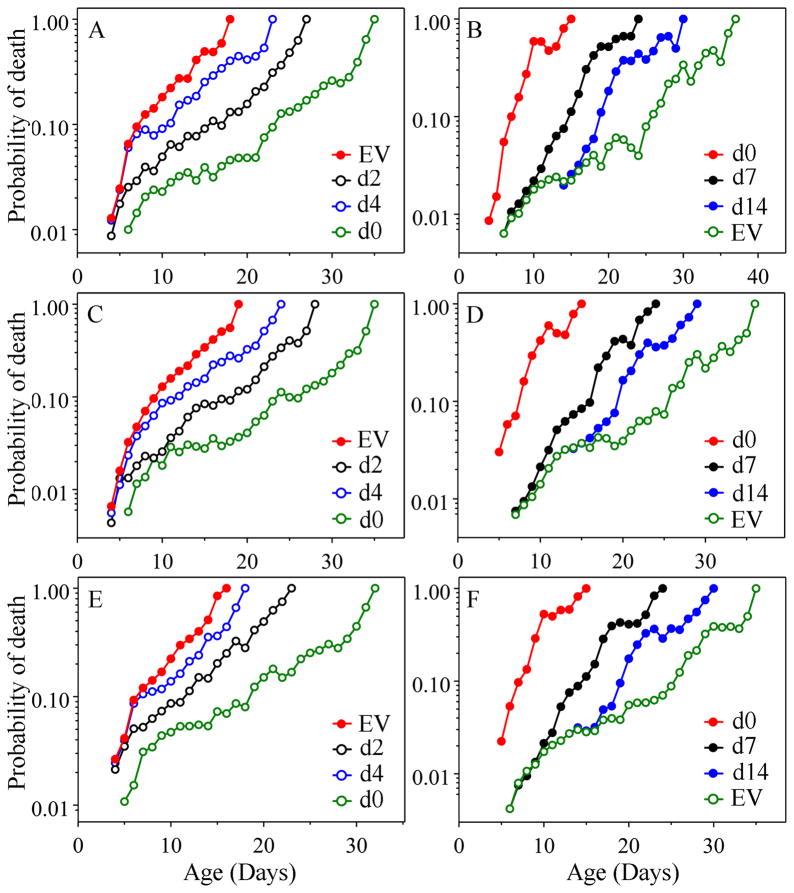
The analysis of age-specific mortality can yield insights into how anti-aging interventions operate that cannot be matched by simple assessment of longevity. Mortality, as opposed to longevity, can be used to assess the effects of an anti-aging intervention on a daily basis, rather than only after most animals have died. Various gerontogene mutations in Caenorhabditis elegans have been shown to increase longevity as much as tenfold and to decrease mortality at some ages even more. Environmental alterations, such as reduced food intake (dietary restriction) and lower temperature also result in reduced mortality soon after the intervention. Here, we ask how soon anti-aging interventions, applied during adult life, affect age-specific mortality in nematodes. Using maximum likelihood analysis, we estimated the Gompertz parameters after shifts of temperature, and of food concentration and maintenance conditions. In separate experiments, we altered expression of age-1 and daf-16, using RNAi. Using about 44 000 nematodes in total, to examine daily mortality, we find that for both types of environmental shift, mortality responded immediately in the first assessment, while RNAi-induced changes resulted in a slower response, perhaps due to delayed mechanics of RNAi action. However, under all conditions there is a permanent 'memory' of past states, such that the initial mortality component [a] of the Gompertz equation [mu(x) = ae(bx)] bears a permanent 'imprint' of that earlier state. However, 'b' (the rate of mortality increase with age) is always specified by the current conditions.
Figures
References
-
- Aballay A, Yorgey P, Ausubel F. Salmonella typhimurium proliferates and establishes a persistent infection in the intestine of Caenorhabditis elegans. Curr Biol. 2000;10:1539–1542. - PubMed
-
- Arantes-Oliveira N, Berman JR, Kenyon C. Healthy animals with extreme longevity. Science. 2003;302:611. - PubMed
-
- Ayyadevara S, Alla R, Thaden JJ, Shmookler Reis RJ. Remarkable longevity and stress resistance of nematode PI3K-null mutants. Aging Cell. 2008;7:13–22. - PubMed
-
- Berrigan D, Perkins SN, Haines DC, Hursting SD. Adult-onset calorie restriction and fasting delay spontaneous tumorigenesis in p53-deficient mice. Carcinogenesis. 2002;23:817–822. - PubMed
-
- Carey JR, Liedo P, Muller HG, Wang JL, Vaupel JW. Dual modes of aging in Mediterranean fruit fly females. Science. 1998;281:996–998. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
