

Hypocretin/orexin in arousal and stress
- PMID: 19748490
- PMCID: PMC2819651
- DOI: 10.1016/j.brainres.2009.09.019
Hypocretin/orexin in arousal and stress
Abstract
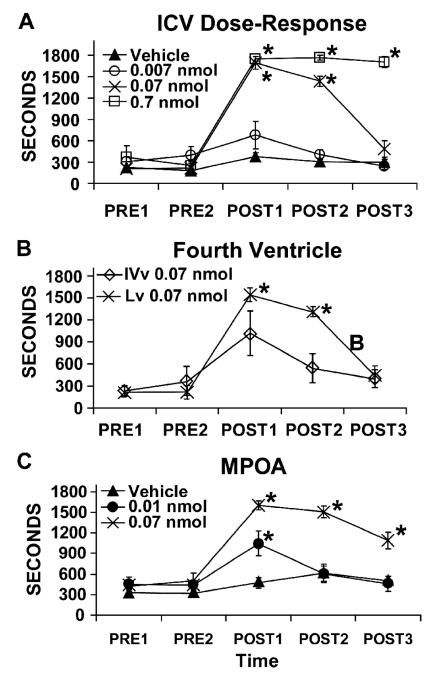
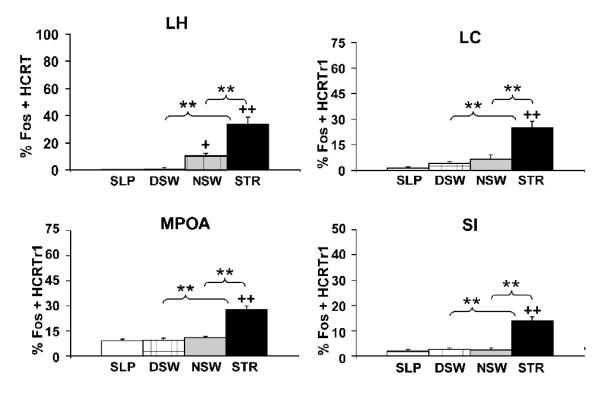
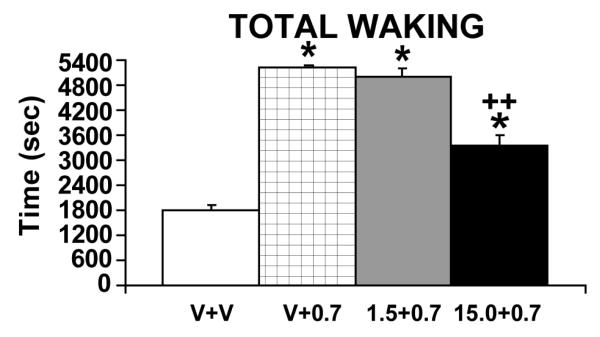
Multiple lines of evidence indicate that hypocretin/orexin (HCRT) participates in the regulation of arousal and arousal-related process. For example, HCRT axons and receptors are found within a variety of arousal-related systems. Moreover, when administered centrally, HCRT exerts robust wake-promoting actions. Finally, a dysregulation of HCRT neurotransmission is associated with the sleep/arousal disorder, narcolepsy. Combined, these observations suggested that HCRT might be a key transmitter system in the regulation of waking. Nonetheless, subsequent evidence indicates that HCRT may not play a prominent role in the initiation of normal waking. Instead HCRT may participate in a variety of processes such as consolidation of waking and/or coupling metabolic state with behavioral state. Additionally, substantial evidence suggests a potential involvement of HCRT in high-arousal conditions, including stress. Thus, HCRT neurotransmission is closely linked to high-arousal conditions, including stress, and HCRT administration exerts a variety of stress-like physiological and behavioral effects that are superimposed on HCRT-induced increases in arousal. Combined, this evidence suggests the hypothesis that HCRT may participate in behavioral responding under high-arousal aversive conditions. Importantly, these actions of HCRT may not be limited to stress. Like stress, appetitive conditions are associated with elevated arousal levels and a stress-like activation of various physiological systems. These and other observations suggest that HCRT may, at least in part, exert affectively neutral actions that are important under high-arousal conditions associated with elevated motivation and/or need for action.
Copyright 2009 Elsevier B.V. All rights reserved.
Figures
References
-
- Al Barazanji KA, Wilson S, Baker J, Jessop DS, Harbuz MS. Central orexin-A activates hHypothalamic-pituitary-adrenal axis and stimulates hypothalamic corticotropin releasing factor and arginine vasopressin neurones in conscious rats. J. Neuroendocrinol. 2001;13:421–424. - PubMed
-
- Antelman SM, Szechtman H, Chin P, Fisher AE. Tail pinch-induced eating, gnawing and licking behavior in rats: dependence on the nigrostriatal dopamine system. Brain Res. 1975;99:319–337. - PubMed
-
- Arnsten AF. Development of the cerebral cortex: XIV. Stress impairs prefrontal cortical function. J. Am. Acad. Child Adolesc. Psychiatry. 1999;38:220–222. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
