

Engagement of TLR2 does not reverse the suppressor function of mouse regulatory T cells, but promotes their survival
- PMID: 19748987
- PMCID: PMC2788511
- DOI: 10.4049/jimmunol.0901465
Engagement of TLR2 does not reverse the suppressor function of mouse regulatory T cells, but promotes their survival
Abstract
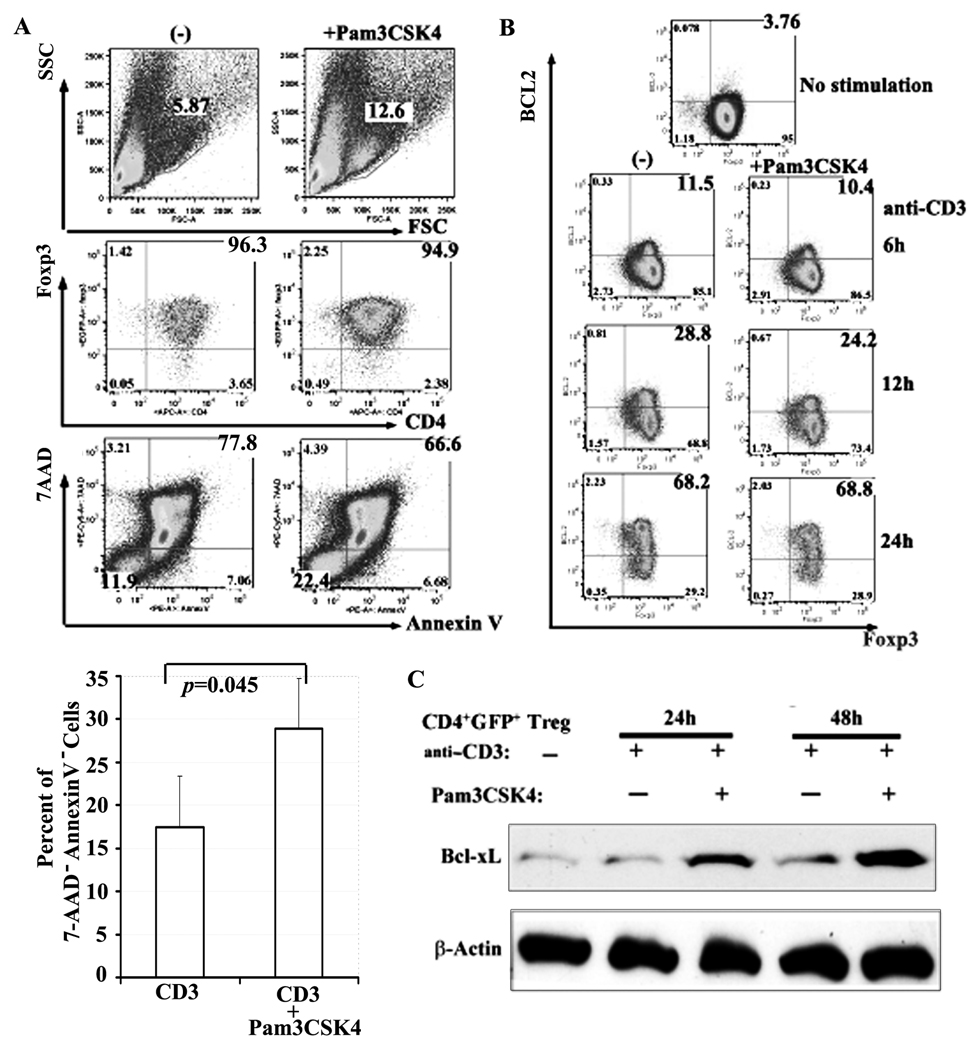
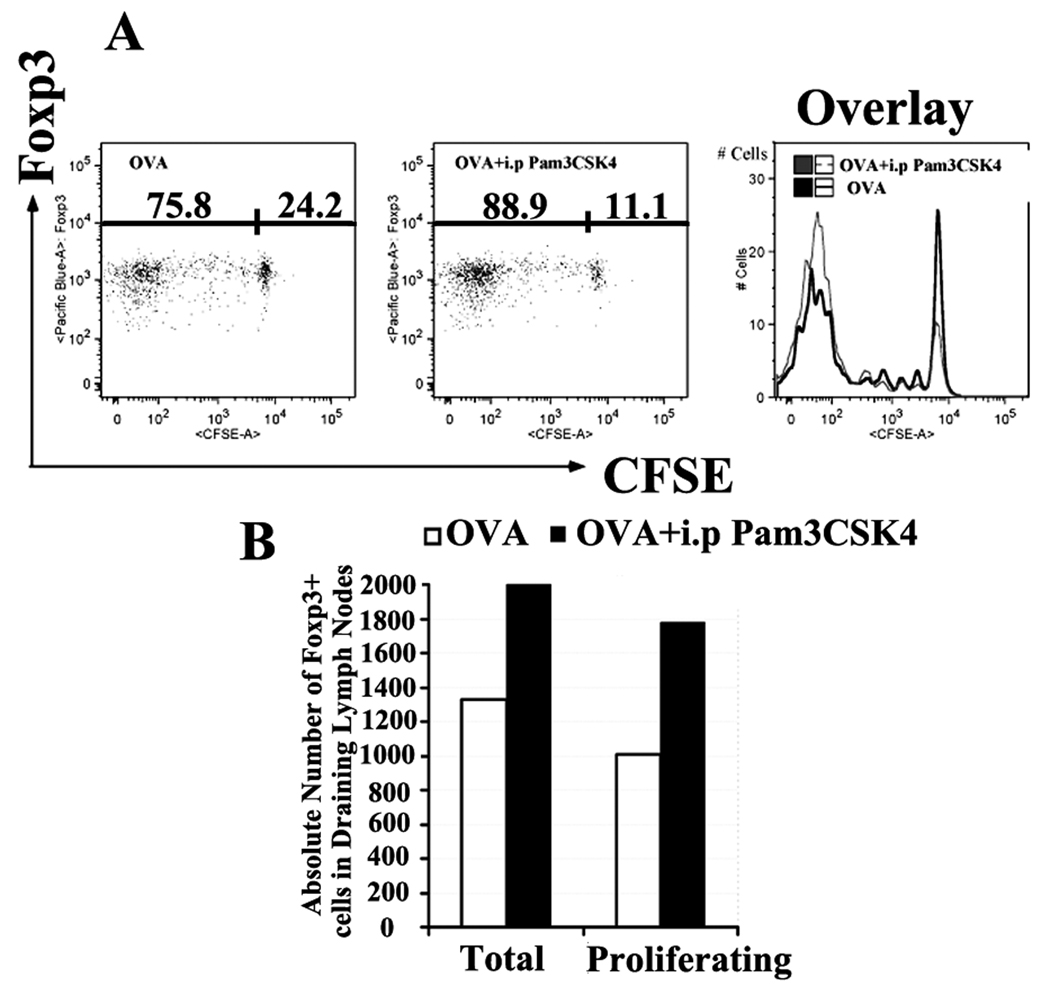
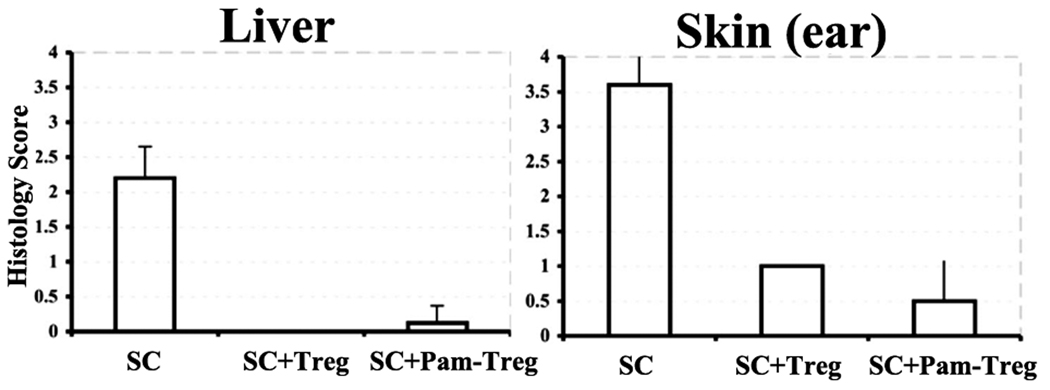
TLRs are a class of conserved pattern recognition receptors that are used by cells of the innate immune system. Recent studies have demonstrated the expression of TLRs on both human and mouse T cells raising the possibility that TLRs play a direct role in adaptive immunity. TLR2 is activated primarily by bacterial wall components including peptidoglycan and lipoproteins. Several studies have shown that mouse regulatory T (Treg) cells express TLR2 and claimed that engagement of TLR2 by synthetic ligands reversed their suppressive function. In contrary, enhancement of Treg function was observed following engagement of TLR2 on human Treg. We have reexamined the expression and function of TLR2 on mouse Treg purified from Foxp3-GFP knock-in mice. TLR2 ligation by TLR2 agonist, the synthetic bacterial lipoprotein Pam3CSK4, enhanced the proliferative responses of both conventional T cells and Treg in response to TLR stimulation in the absence of APC. Treatment of Foxp3+ Treg with Pam3CSK4 did not alter their suppressive function in vitro or in vivo and did not reduce their level of Foxp3 expression. An additional effect of TLR2 stimulation of Treg was induction of Bcl-x(L) resulting in enhanced survival in vitro. Treatment of mice with the TLR2 agonist enhanced the Ag-driven proliferation of Treg in vivo, but did not abolish their ability to suppress the development of experimental autoimmune encephalomyelitis. Development of methods to selectively stimulate TLR2 on Treg may lead to a novel approaches for the treatment of autoimmune diseases.
Conflict of interest statement
The authors have no financial conflict of interest.
Figures





References
-
- Takeda K, Akira S. Toll receptors and pathogen resistance. Cell. Microbiol. 2003;5:143–153. - PubMed
-
- Takeda K, Kaisho T, Akira S. Toll-like receptors. Annu. Rev. Immunol. 2003;21:335–376. - PubMed
-
- Kabelitz D. Expression and function of Toll-like receptors in T lymphocytes. Curr. Opin. Immunol. 2007;19:39–45. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
