

doi: 10.1128/JB.01000-09.
Epub 2009 Sep 11.
YjhS (NanS) is required for Escherichia coli to grow on 9-O-acetylated N-acetylneuraminic acid
Affiliations
- PMID: 19749043
- PMCID: PMC2772488
- DOI: 10.1128/JB.01000-09
Item in Clipboard
YjhS (NanS) is required for Escherichia coli to grow on 9-O-acetylated N-acetylneuraminic acid
J Bacteriol.
2009 Nov.
Abstract
The nanATEK-yhcH, yjhATS, and yjhBC operons in Escherichia coli are coregulated by environmental N-acetylneuraminic acid, the most prevalent sialic acid in nature. Here we show that YjhS (NanS) is a probable 9-O-acetyl N-acetylneuraminic acid esterase required for E. coli to grow on this alternative sialic acid, which is commonly found in mammalian host mucosal sites.
Figures
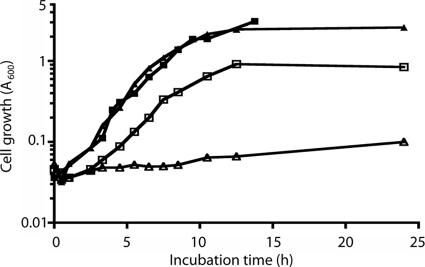
nanS is required for E. coli to grow on Neu5,9Ac2 as the sole carbon source. Overnight cultures of BW30270 and its isogenic nanS mutant derivative grown on glycerol as the carbon source were diluted 1:50 into fresh medium containing either glycerol or Neu5,9Ac2 as follows: BW30270 with glycerol (closed squares); nanS mutant with glycerol (closed triangles); BW30270 with Neu5,9Ac2 (open squares); nanS mutant with Neu5,9Ac2 (open triangles).
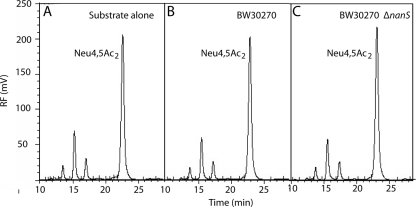
Chemical analysis of Neu4,5Ac2 in spent culture medium. (A) Neu4,5Ac2 at a 0.1% final concentration was incubated in minimal glycerol medium without cells for 7 h and then subjected to DMB analysis as described in the text. (B) Neu4,5Ac2 in the spent culture supernatant of wild type was grown for 7 h in the presence of glycerol. (C) Neu4,5Ac2 in the spent culture supernatant of the nanS mutant was grown in glycerol medium for 7 h. Note that the same profiles were obtained after comparable chemical analyses of 24-h cultures, indicating the stability of Neu4,5Ac2 and its resistance to metabolism by E. coli K-12. RF, relative fluorescence.
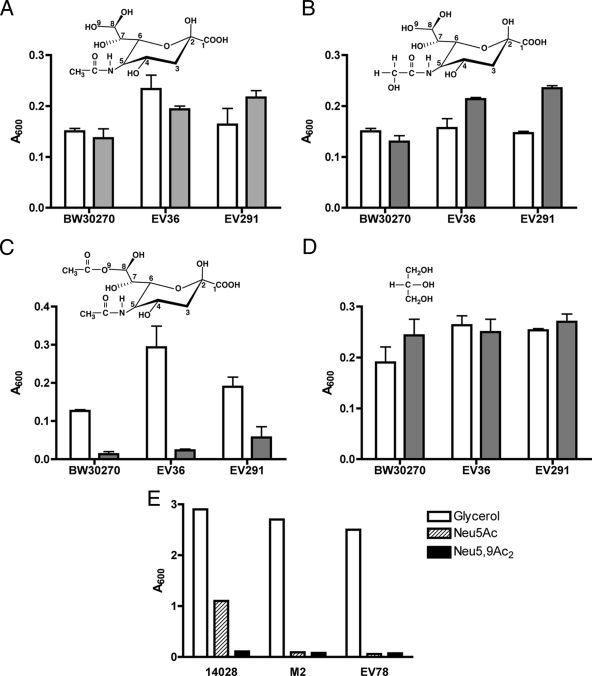
Growth of the wild type and the yjhS and nanA mutants on different sialic acids. In panels A to D, the indicated wild-type strains (open boxes) and their isogenic yjhS deletion derivatives (shaded boxes) were diluted 50-fold from stationary-phase cells grown in M63 medium plus glycerol (0.4%) into 0.1 ml of M63 medium containing 0.1% of the indicated sialic acids or glycerol in 96-well plates. (A) Growth on Neu5Ac. (B) Growth on Neu5Gc. (C) Growth on Neu5,9Ac2. (D) Growth on glycerol. The plate was incubated without shaking at 37°C for 16 h, and A600 values were recorded with a microplate reader. Data represent the means of three independent experiments ± standard deviations. (E) S. enterica serovar Typhimurium wild-type strain 14028, nanA strain LT2 derivative M2, and E. coli nanA mutant EV78 were diluted 60-fold into 0.5 ml of M63 medium containing 0.4% glycerol, 0.1% Neu5Ac, or 0.1% Neu5,9Ac2. Cultures were grown at 37°C with vigorous aeration; the A600 values were recorded after 7 h.
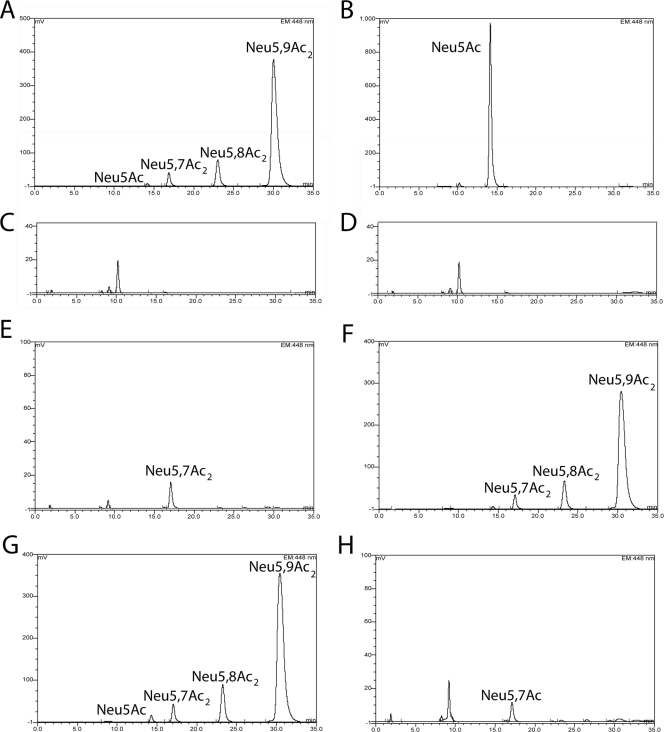
Utilization of Neu5Ac or Neu5,9Ac2 by BW30270 and its yjhS deletion derivative growing simultaneously in the presence of glycerol. BW30270 or its yjhS derivative was diluted 60-fold into 0.5 ml of M63 medium plus glycerol containing either 0.1% Neu5Ac or Neu5,9Ac2 and incubated at 37°C with vigorous aeration for 7 h unless indicated otherwise. (A) Neu5,9Ac2 incubated without cells. (B) Neu5Ac incubated without cells. (C) Wild type grown with Neu5Ac. (D) Mutant grown with Neu5Ac. Note that the minor peak eluting at about 10 min as shown in these panels and in the succeeding panel is likely to represent a trace amount of 2-deoxy-d -manno-octulosonic acid derived from sloughed lipopolysaccharide during the incubation that was not removed by the centrifugation step used to produce the spent culture media for the analyses (22). (E) Wild type grown with Neu5,9Ac2. (F) Mutant grown with Neu5,9Ac2. (G) Neu5,9Ac2 incubated without cells for 17 h. (H) Mutant grown with Neu5,9Ac2 for 17 h.
Similar articles
-
Unexpected Diversity of Escherichia coli Sialate O-Acetyl Esterase NanS.J Bacteriol. 2016 Sep 22;198(20):2803-9. doi: 10.1128/JB.00189-16. Print 2016 Oct 15. J Bacteriol. 2016. PMID: 27481927 Free PMC article.
-
Control of the Escherichia coli sialoregulon by transcriptional repressor NanR.J Bacteriol. 2013 Oct;195(20):4689-701. doi: 10.1128/JB.00692-13. Epub 2013 Aug 9. J Bacteriol. 2013. PMID: 23935044 Free PMC article.
-
De-O-Acetylation of mucin-derived sialic acids by recombinant NanS-p esterases of Escherichia coli O157:H7 strain EDL933.Int J Med Microbiol. 2018 Dec;308(8):1113-1120. doi: 10.1016/j.ijmm.2018.10.001. Epub 2018 Oct 9. Int J Med Microbiol. 2018. PMID: 30340996 Free PMC article.
-
Diversity of microbial sialic acid metabolism.Microbiol Mol Biol Rev. 2004 Mar;68(1):132-53. doi: 10.1128/MMBR.68.1.132-153.2004. Microbiol Mol Biol Rev. 2004. PMID: 15007099 Free PMC article. Review.
-
Sialic acid diversity in the human gut: Molecular impacts and tools for future discovery.Curr Opin Struct Biol. 2022 Aug;75:102397. doi: 10.1016/j.sbi.2022.102397. Epub 2022 May 30. Curr Opin Struct Biol. 2022. PMID: 35653953 Review.
Cited by
-
Role of sialidase in glycoprotein utilization by Tannerella forsythia.Microbiology (Reading). 2011 Nov;157(Pt 11):3195-3202. doi: 10.1099/mic.0.052498-0. Epub 2011 Sep 1. Microbiology (Reading). 2011. PMID: 21885482 Free PMC article.
-
Growth media simulating ileal and colonic environments affect the intracellular proteome and carbon fluxes of enterohemorrhagic Escherichia coli O157:H7 strain EDL933.Appl Environ Microbiol. 2013 Jun;79(12):3703-15. doi: 10.1128/AEM.00062-13. Epub 2013 Apr 5. Appl Environ Microbiol. 2013. PMID: 23563955 Free PMC article.
-
Social-Stress-Responsive Microbiota Induces Stimulation of Self-Reactive Effector T Helper Cells.mSystems. 2019 May 14;4(4):e00292-18. doi: 10.1128/mSystems.00292-18. eCollection 2019 Jul-Aug. mSystems. 2019. PMID: 31098398 Free PMC article.
-
"Just a spoonful of sugar...": import of sialic acid across bacterial cell membranes.Biophys Rev. 2018 Apr;10(2):219-227. doi: 10.1007/s12551-017-0343-x. Epub 2017 Dec 8. Biophys Rev. 2018. PMID: 29222808 Free PMC article. Review.
-
Sialic acid, periodontal pathogens and Tannerella forsythia: stick around and enjoy the feast!Mol Oral Microbiol. 2012 Feb;27(1):11-22. doi: 10.1111/j.2041-1014.2011.00630.x. Epub 2011 Nov 12. Mol Oral Microbiol. 2012. PMID: 22230462 Free PMC article. Review.
References
-
- Baba, T., T. Ara, M. Hasegawa, Y. Takai, Y. Okumura, M. Baba, K. A. Datsenko, M. Tomita, B. L. Wanner, and H. Mori. 21 February 2006, posting date. Construction of Escherichia coli K-12 in-frame, single-gene knockout mutants: the Keio collection. Mol. Syst. Biol. doi:10.1038/msb4100050. - DOI - PMC - PubMed
-
- Becker, D., M. Selbach, C. Rollenhagen, M. Ballmaier, T. F. Meyer, M. Mann, and D. Bumann. 2006. Robust Salmonella metabolism limits possibilities for new antimicrobials. Nature 440:303-307. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
