

Decorin core protein (decoron) shape complements collagen fibril surface structure and mediates its binding
- PMID: 19753304
- PMCID: PMC2737631
- DOI: 10.1371/journal.pone.0007028
Decorin core protein (decoron) shape complements collagen fibril surface structure and mediates its binding
Abstract
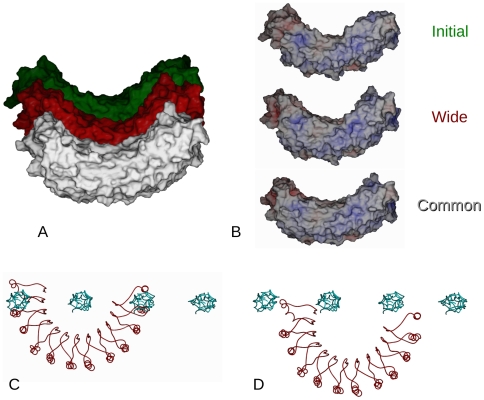
Decorin is the archetypal small leucine rich repeat proteoglycan of the vertebrate extracellular matrix (ECM). With its glycosaminoglycuronan chain, it is responsible for stabilizing inter-fibrillar organization. Type I collagen is the predominant member of the fibrillar collagen family, fulfilling both organizational and structural roles in animal ECMs. In this study, interactions between decoron (the decorin core protein) and binding sites in the d and e(1) bands of the type I collagen fibril were investigated through molecular modeling of their respective X-ray diffraction structures. Previously, it was proposed that a model-based, highly curved concave decoron interacts with a single collagen molecule, which would form extensive van der Waals contacts and give rise to strong non-specific binding. However, the large well-ordered aggregate that is the collagen fibril places significant restraints on modes of ligand binding and necessitates multi-collagen molecular contacts. We present here a relatively high-resolution model of the decoron-fibril collagen complex. We find that the respective crystal structures complement each other well, although it is the monomeric form of decoron that shows the most appropriate shape complementarity with the fibril surface and favorable calculated energies of interaction. One molecule of decoron interacts with four to six collagen molecules, and the binding specificity relies on a large number of hydrogen bonds and electrostatic interactions, primarily with the collagen motifs KXGDRGE and AKGDRGE (d and e(1) bands). This work helps us to understand collagen-decorin interactions and the molecular architecture of the fibrillar ECM in health and disease.
Conflict of interest statement
Figures





References
-
- Daley W, Peters S, Larsen M. Extracellular matrix dynamics in development and regenerative medicine. Journal of cell science. 2008;121:255–64. - PubMed
-
- Scott JE. Supramolecular organization of extracellular matrix glycosaminoglycans, in vitro and in the tissues. The FASEB journal. 1992;6:2639–45. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
