

The alphabeta T cell receptor is an anisotropic mechanosensor
- PMID: 19755427
- PMCID: PMC2781503
- DOI: 10.1074/jbc.M109.052712
The alphabeta T cell receptor is an anisotropic mechanosensor
Abstract
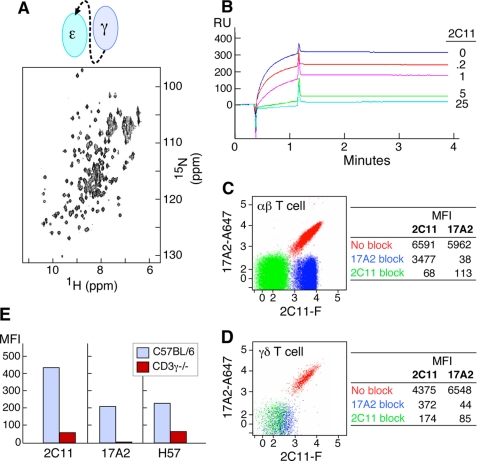
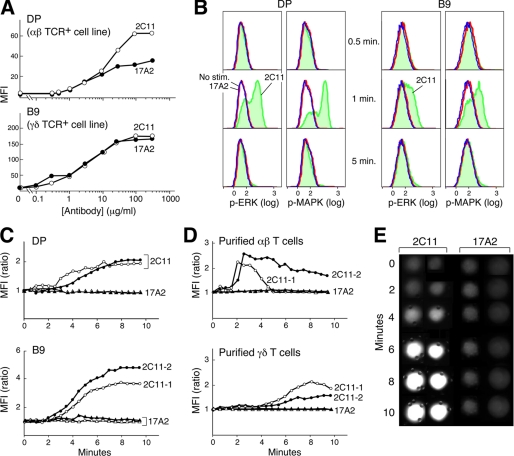
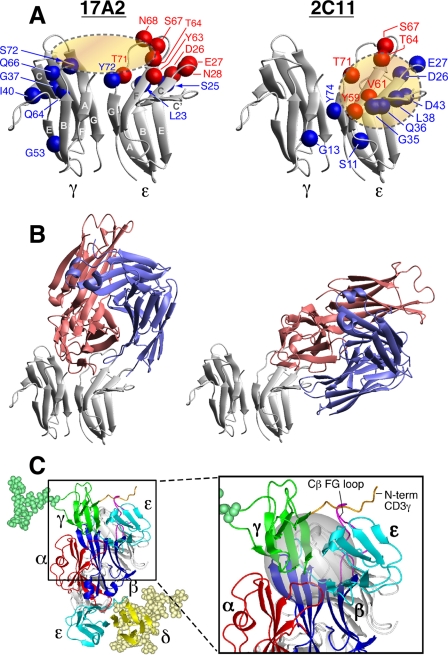
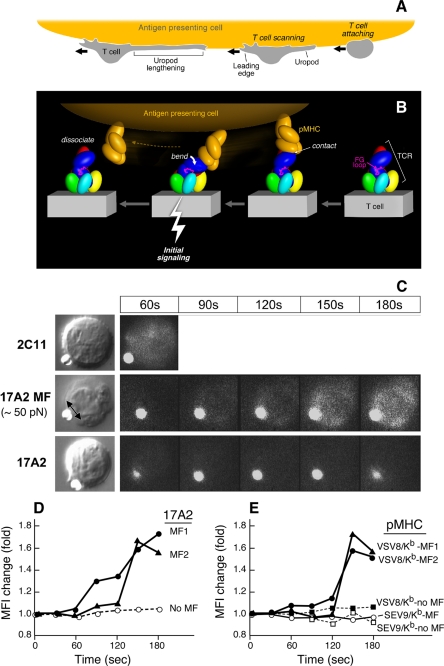
Thymus-derived lymphocytes protect mammalian hosts against virus- or cancer-related cellular alterations through immune surveillance, eliminating diseased cells. In this process, T cell receptors (TCRs) mediate both recognition and T cell activation via their dimeric alphabeta, CD3 epsilon gamma, CD3 epsilon delta, and CD3 zeta zeta subunits using an unknown structural mechanism. Here, site-specific binding topology of anti-CD3 monoclonal antibodies (mAbs) and dynamic TCR quaternary change provide key clues. Agonist mAbs footprint to the membrane distal CD3 epsilon lobe that they approach diagonally, adjacent to the lever-like C beta FG loop that facilitates antigen (pMHC)-triggered activation. In contrast, a non-agonist mAb binds to the cleft between CD3 epsilon and CD3 gamma in a perpendicular mode and is stimulatory only subsequent to an external tangential but not a normal force ( approximately 50 piconewtons) applied via optical tweezers. Specific pMHC but not irrelevant pMHC activates a T cell upon application of a similar force. These findings suggest that the TCR is an anisotropic mechanosensor, converting mechanical energy into a biochemical signal upon specific pMHC ligation during immune surveillance. Activating anti-CD3 mAbs mimic this force via their intrinsic binding mode. A common TCR quaternary change rather than conformational alterations can better facilitate structural signal initiation, given the vast array of TCRs and their specific pMHC ligands.
Figures
References
-
- Rudolph M. G., Stanfield R. L., Wilson I. A. (2006) Annu. Rev. Immunol. 24, 419–466 - PubMed
-
- Iwashima M., Irving B. A., van Oers N. S., Chan A. C., Weiss A. (1994) Science 263, 1136–1139 - PubMed
-
- Shinkai Y., Ma A., Cheng H. L., Alt F. W. (1995) Immunity 2, 401–411 - PubMed
-
- Sprent J., Surh C. D. (2002) Annu. Rev. Immunol. 20, 551–579 - PubMed
-
- Teixeiro E., Daniels M. A., Hamilton S. E., Schrum A. G., Bragado R., Jameson S. C., Palmer E. (2009) Science 323, 502–505 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
