

Review
doi: 10.1038/nrmicro2204.
Cholera transmission: the host, pathogen and bacteriophage dynamic
Affiliations
- PMID: 19756008
- PMCID: PMC3842031
- DOI: 10.1038/nrmicro2204
Item in Clipboard
Review
Cholera transmission: the host, pathogen and bacteriophage dynamic
Nat Rev Microbiol.
2009 Oct.
Abstract
Zimbabwe offers the most recent example of the tragedy that befalls a country and its people when cholera strikes. The 2008-2009 outbreak rapidly spread across every province and brought rates of mortality similar to those witnessed as a consequence of cholera infections a hundred years ago. In this Review we highlight the advances that will help to unravel how interactions between the host, the bacterial pathogen and the lytic bacteriophage might propel and quench cholera outbreaks in endemic settings and in emergent epidemic regions such as Zimbabwe.
Figures
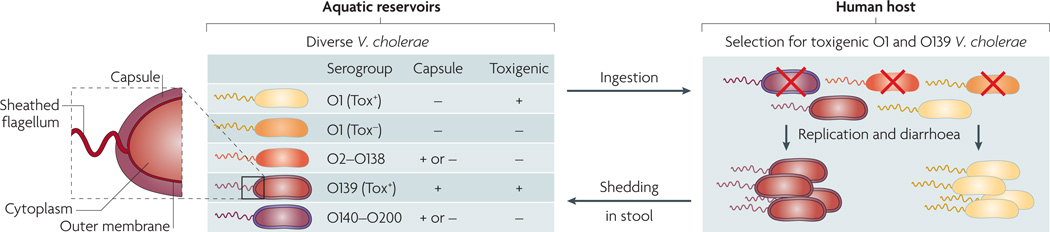
On the basis of the antigenicity of the O antigen component of the outer membrane lipopolysaccharide, more than 200 serogroups (O1–O200) of Vibrio cholerae exist in aquatic environments. Only a subset of O1 and O139 serogroup strains are toxigenic (Tox+) and therefore capable of causing cholera when ingested; such strains are selected for in the host. Other strains are non-toxigenic (Tox−) and are selected against. Different O antigen types are indicated by the colour of the outer membrane and sheathed flagellum (the periplasmic space and the inner membrane are not shown). Capsules are present in a subset of strains. Different strain genotypes are indicated by the colour of the cytoplasm; note that Tox+ O1 and O139 have essentially the same genotype, with the exception of the O antigen genes.
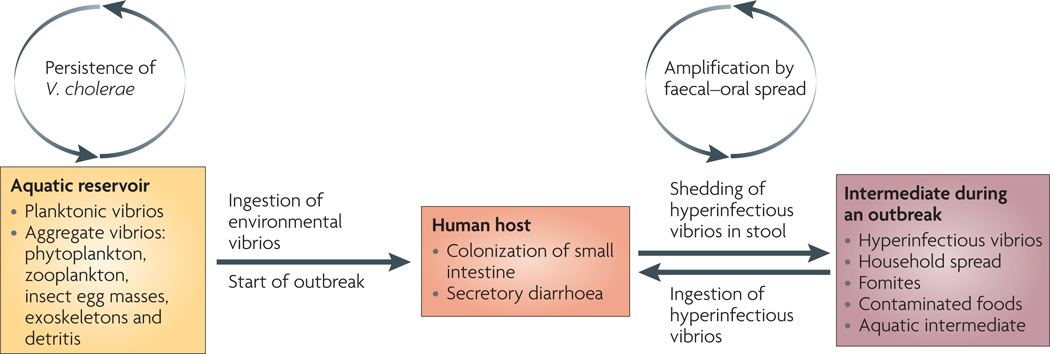
Toxigenic strains of Vibrio cholerae persist in aquatic environments alongside non-toxigenic strains, aided by biofilm formation on biological surfaces and use of chitin as a carbon and nitrogen source. On ingestion of these aquatic-environment-adapted bacteria in contaminated food or water, toxigenic strains colonize the small intestine, multiply, secrete cholera toxin and are shed back into the environment by the host in secretory diarrhoea. The stool-shed pathogens are in a transient hyperinfectious state that serves to amplify the outbreak through transmission to subsequent hosts.
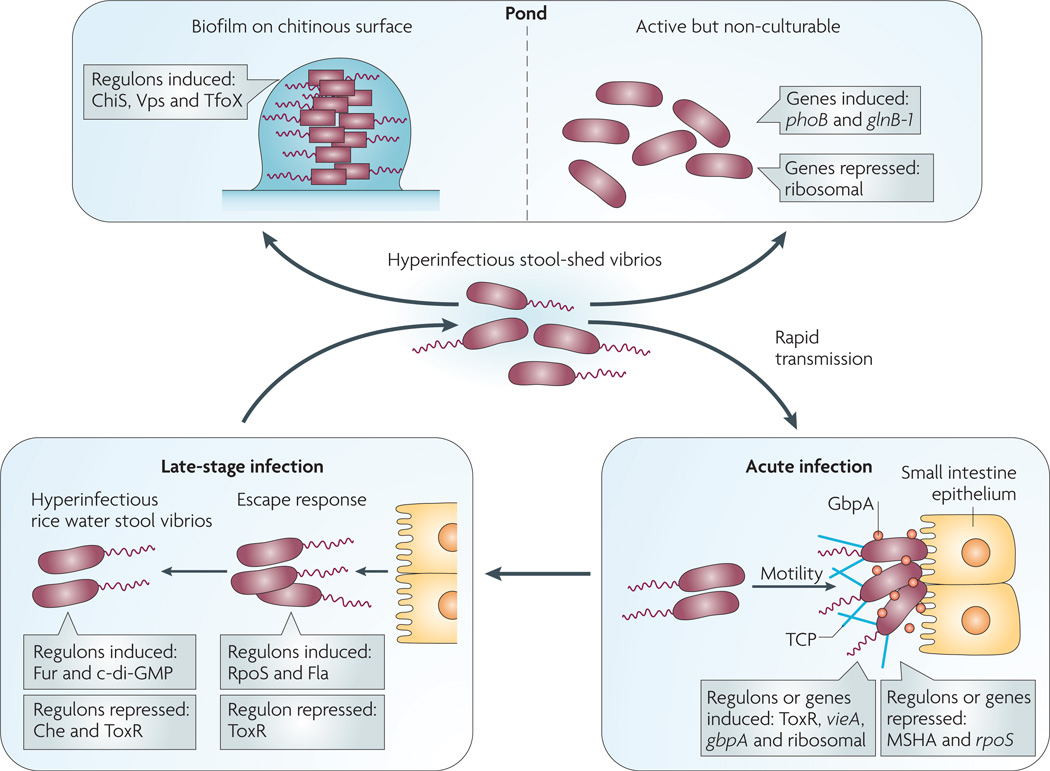
On ingestion (bottom right panel), Vibrio cholerae uses motility and mucinases to penetrate the mucus gel and N-acetylglucosamine-binding protein A (GbpA ) and other factors to colonize the small intestine epithelium. Substantial changes in gene expression accompany this transition from pond to acute infection including, but not limited to, induction of vieA , which encodes a phosphodiesterase that hydrolyzes the second messenger cyclic di-GMP, and ToxR -regulated genes, including those for cholera toxin and the self-aggregative toxin-coregulated pilus (TCP). In addition several genes are repressed, such as those for the chitin-binding mannose-sensitive hemagglutinin pilus (MSHA) and the stress sigma factor RpoS. At a late stage of infection (bottom left panel), V. cholerae alters its gene expression again in order to detach from the epithelium — the ‘escape response’ — and to prepare for transmission to another host (for example, household spread) or entry into the aquatic environment. The late-stage changes include induction of genes for c-di-GMP synthesis (diguanylyl cyclases), nutrient-scavenging systems (such as the ferric uptake regulation (Fur) regulon) and motility (Fla) as well as repression of genes such as those for chemotaxis (Che) and the ToxR regulon. The resulting ‘motile but non-chemotactic’ state contributes to hyperinfectivity. If bacteria shed in stool are not ingested by another host in a short period of time, then one of two fates awaits them (top panel): establishment in the aquatic environment by finding suitable nutrient sources such as chitin, or decay into an ‘active but non-culturable’ state. Upon exposure to chitin, V. cholerae induces several genes involved in adherence to and catabolism of chitin (the ChiS regulon), as well as those involved in genetic competence (the TfoX regulon). Biofilm formation on surfaces is mediated by induction of the Vps regulon, which encodes extracellular polysaccharide. During transition to the active but non-culturable state, large changes in gene expression occur as V. cholerae attempts to adapt to nutrient-poor conditions. These include induction of phosphate and nitrogen starvation genes (phoB and glnB-1 , respectively) and repression of translation machinery genes.
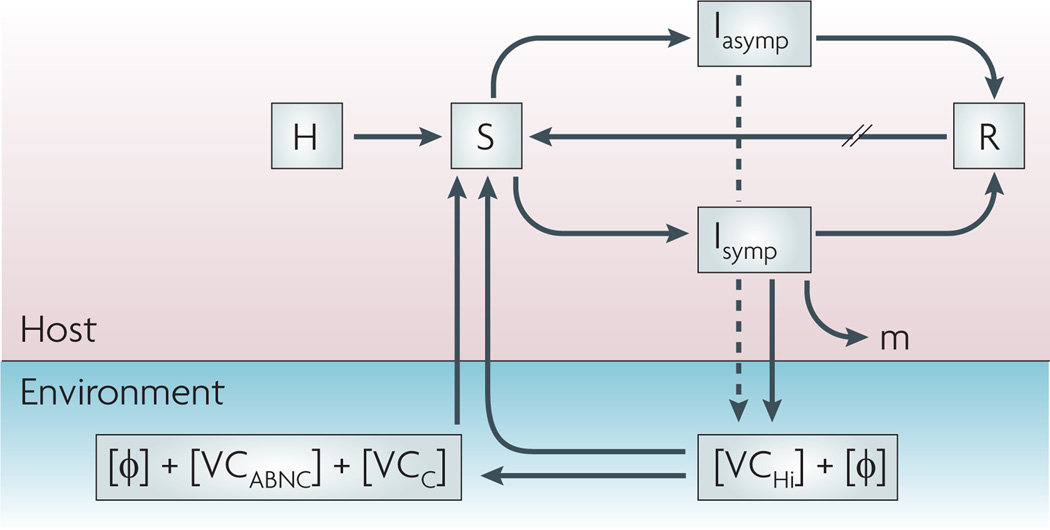
The total population (H) feeds the pool of susceptible hosts (S) that become infectious (I) after consuming Vibrio cholerae from an environmental source, with or without lytic bacteriophages (Φ). Infected individuals are symptomatic (Isymp) or asymptomatic (Iasymp) and recover (R) through the actions of their immune systems and possibly those of lytic bacteriophages, or succumb to the infection (m). Recovered individuals will re-enter the susceptible pool at varying rates (//) depending on the degree of protective immunity. Lytic phages and hyperinfectious V. cholerae(VCHi) are shed by the symptomatic host in varying concentrations; asymptomatic hosts shed far fewer bacteria (dashed line). VCHi cells rapidly passage to the next host, persist in the environment as culturable cells with unknown infectivity (VCC) or decay into an ‘active but non-culturable’ state (VCABNC) with reduced infectivity. All three cell types plus aggregate bacteria (not shown) probably have mixed roles as environmental reservoirs for future outbreaks.
References
-
- Bryce J, Boschi-Pinto C, Shibuya K, Black RE. WHO estimates of the causes of death in children. Lancet. 2005;365:1147–1152. - PubMed
-
- Sack DA, Sack RB, Chaignat CL. Getting serious about cholera. N. Engl. J. Med. 2006;355:649–651. - PubMed
-
- Pollitzer R, Swaroop S, Burrows W. Cholera. Monogr. Ser. World Health Organ. 1959;58:1001–1019. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
