

GB virus type C interactions with HIV: the role of envelope glycoproteins
- PMID: 19758271
- PMCID: PMC3543829
- DOI: 10.1111/j.1365-2893.2009.01194.x
GB virus type C interactions with HIV: the role of envelope glycoproteins
Abstract
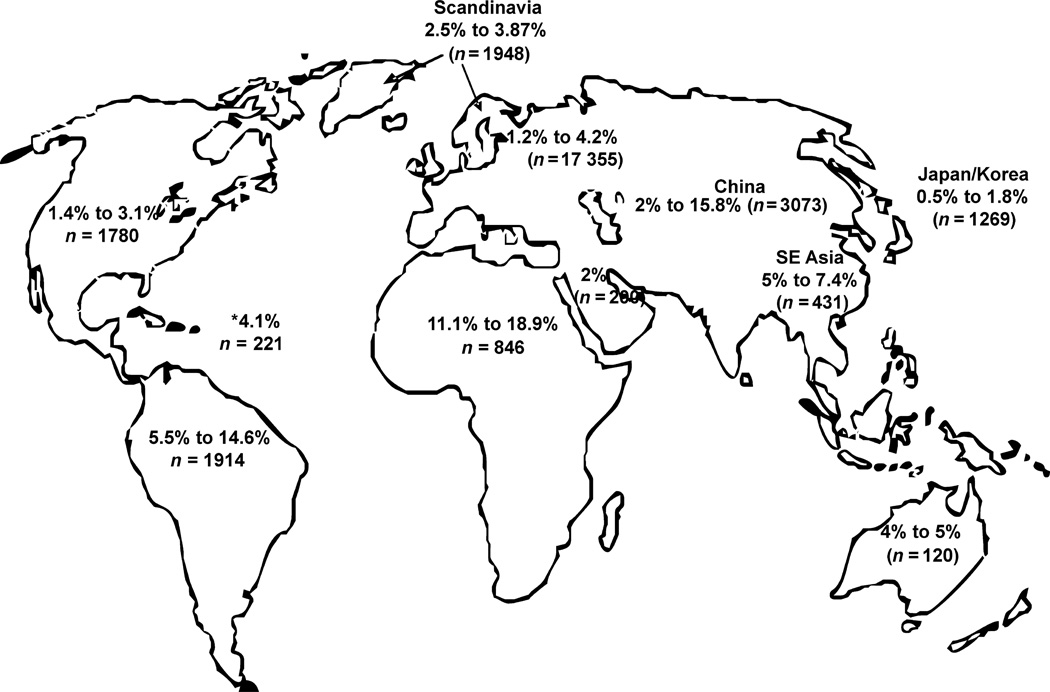
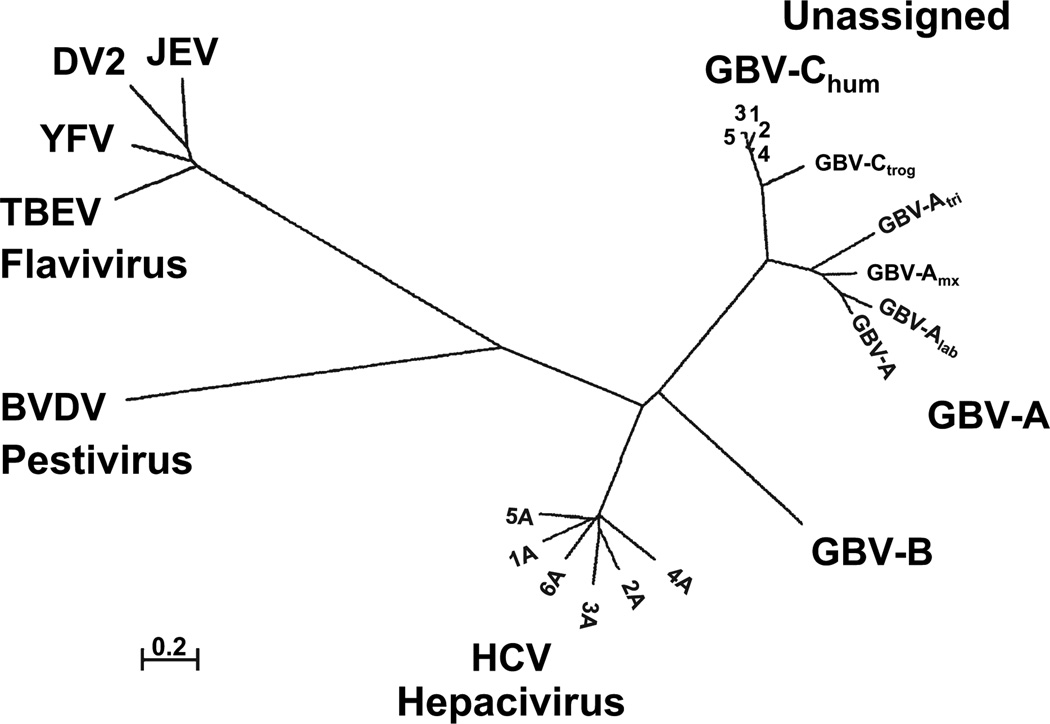
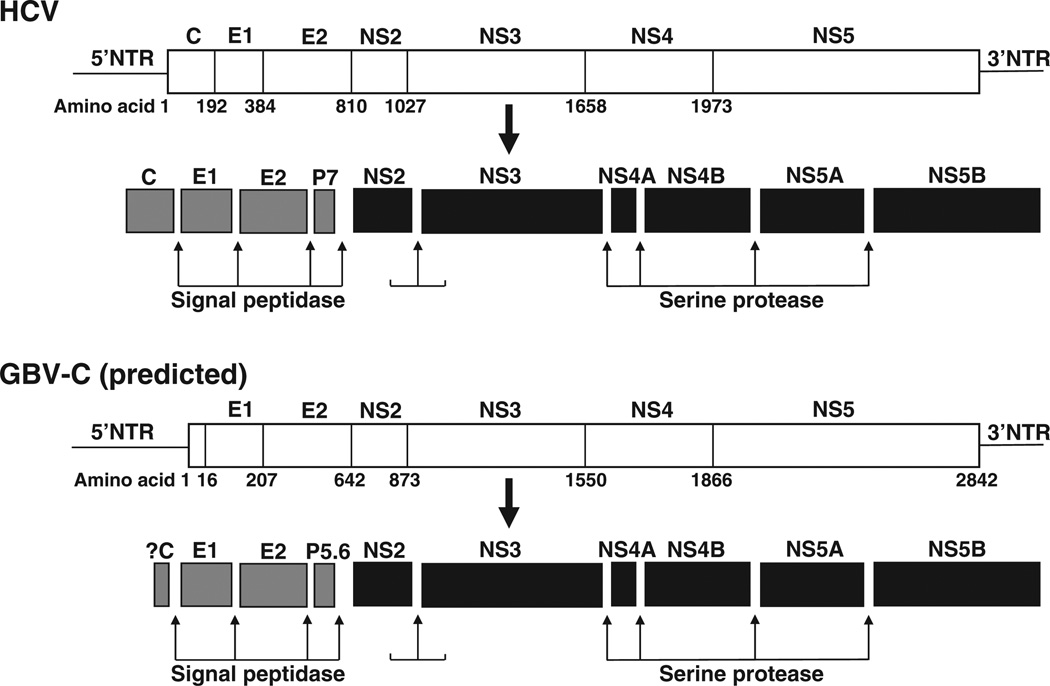
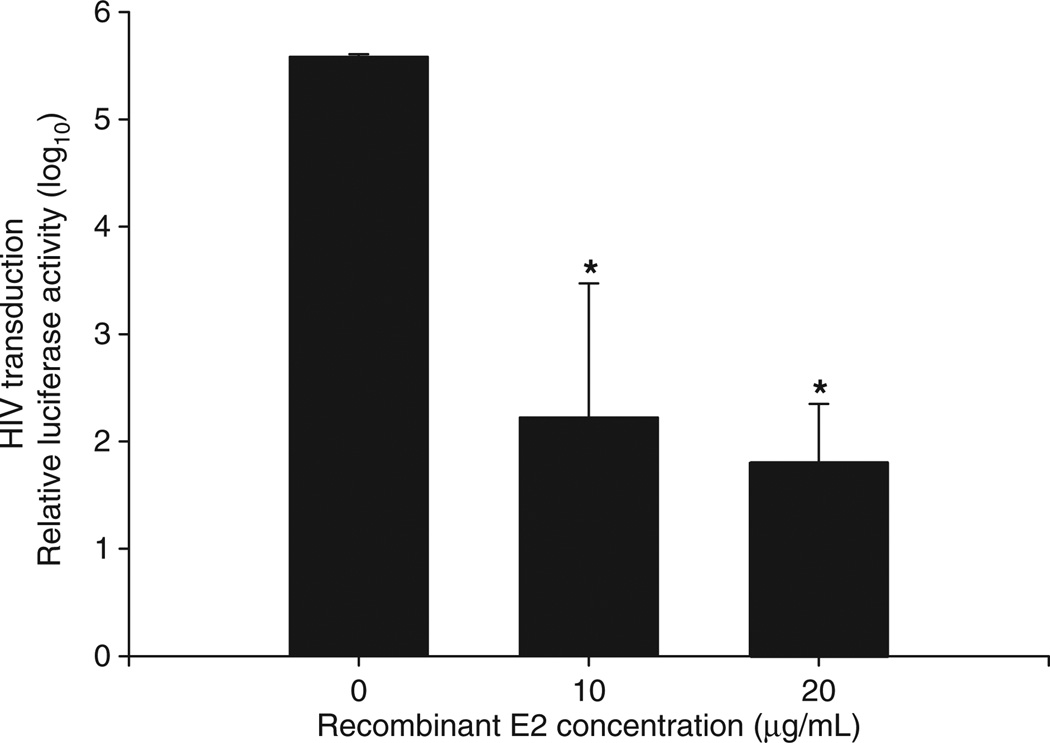
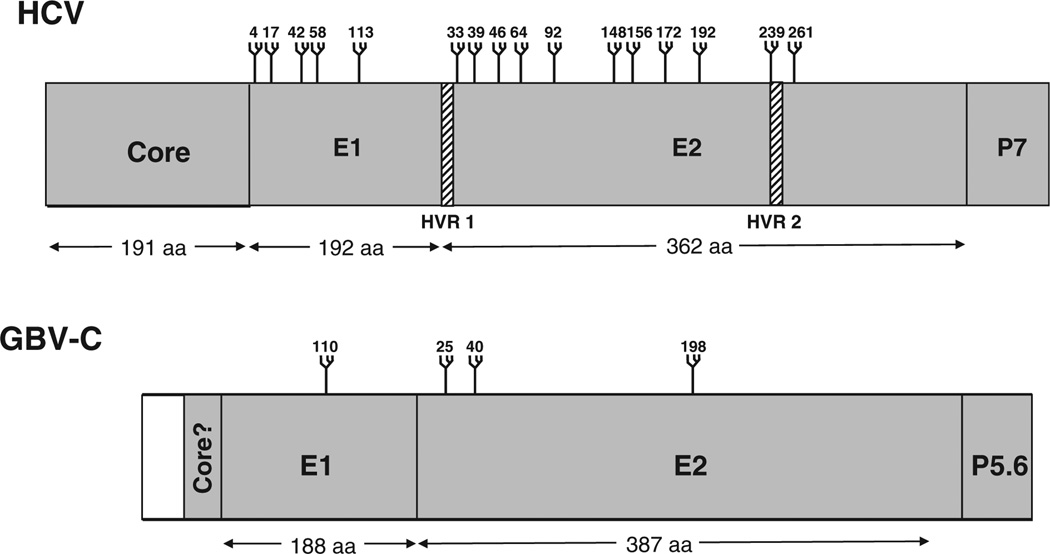
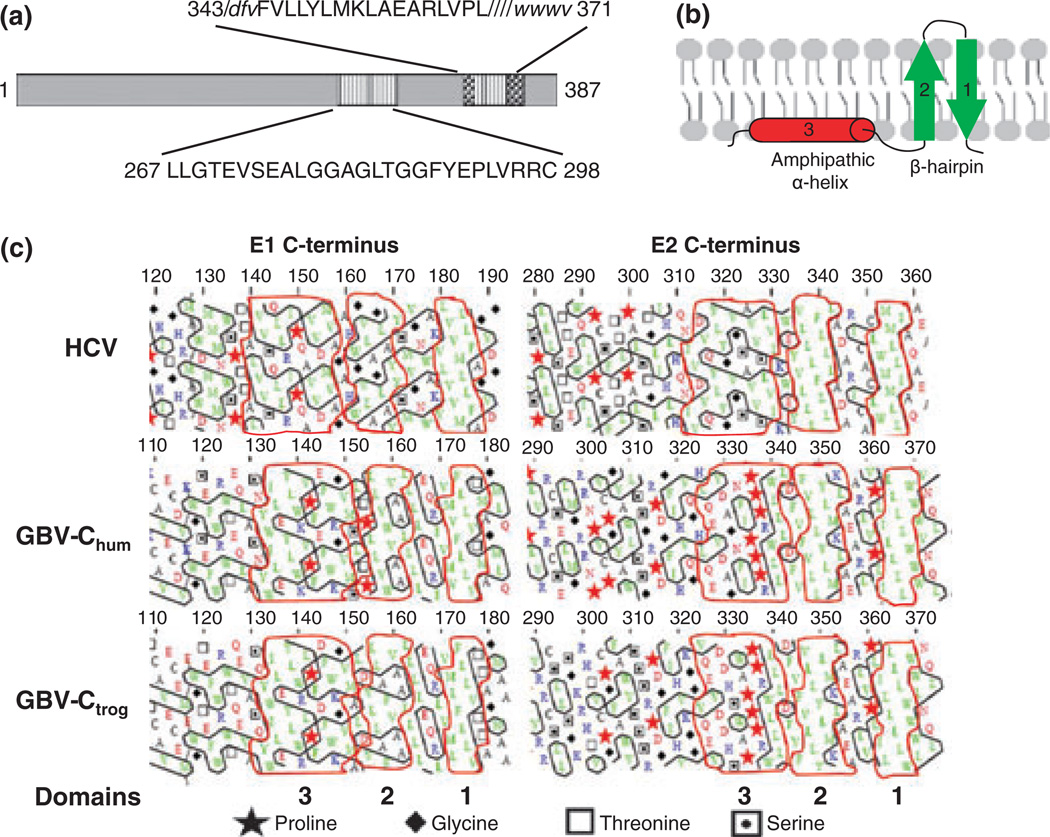
GB virus C/hepatitis G virus (GBV-C/HGV) is the most closely related human virus to hepatitis C virus (HCV). GBV-C is lymphotropic and not associated with any known disease, although it is associated with improved survival in HIV-infected individuals. In peripheral blood mononuclear cells, GBV-C induces the release of soluble ligands for HIV entry receptors (RANTES, MIP-1a, MIP-1b and SDF-1), suggesting that GBV-C may interact with lymphocytes to induce a chemokine and/or cytokine milieu that is inhibitory to HIV infection. Expression of GBV-C envelope glycoprotein E2 in CD4+ T cells or addition of recombinant E2 to CD4 cells recapitulates the HIV inhibition seen with GBV-C infection. Like HCV E2, GBV-C E2 is predicted to be post-translationally processed in the endoplasmic reticulum and is involved with cell binding. The C-termini of GBV-C E1 and E2 proteins contain predicted transmembrane domains sharing features with HCV TM domains. To date, cellular receptor(s) for GBV-C E2 have not been identified. GBV-C E2-mediated HIV inhibition is dose-dependent and HIV replication is blocked at the binding and/or entry step. In addition, a putative GBV-C E2 fusion peptide interferes with HIV gp41 peptide oligomerization required for HIV-1 fusion, further suggesting that GBV-C E2 may inhibit HIV entry. Additional work is needed to identify the GBV-C E2 cellular receptor, characterize GBV-C E2 domains responsible for HIV inhibition, and to examine GBV-C E2-mediated fusion in the context of the entire envelope protein or viral-particles. Understanding the mechanisms of action may identify novel approaches to HIV therapy.
Figures
References
-
- Simons JN, Leary TP, Dawson GJ, et al. Isolation of novel virus-like sequences associated with human hepatitis. Nat Med. 1995;1:564–569. - PubMed
-
- Linnen J, Wages J, Zhang-Keck Z-Y, et al. Molecular cloning and disease association of hepatitis G virus: a transfusion-transmissible agent. Science. 1996;271:505–508. - PubMed
-
- Schaluder GG, Dawson GJ, Simons JN, et al. Molecular and serologic analysis in the transmission of the GB hepatitis agents. J Med Virol. 1995;46:81–90. - PubMed
-
- Alter HJ. G-pers creepers, where’d you get those papers? A reassessment of the literature on the hepatitis G virus. Transfusion. 1997;37:569–572. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
