

Association of the pr peptides with dengue virus at acidic pH blocks membrane fusion
- PMID: 19759134
- PMCID: PMC2786737
- DOI: 10.1128/JVI.01637-09
Association of the pr peptides with dengue virus at acidic pH blocks membrane fusion
Abstract
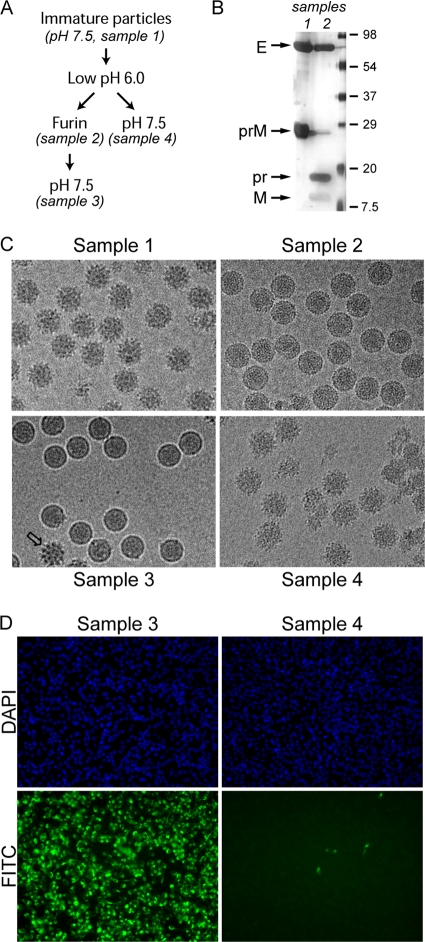
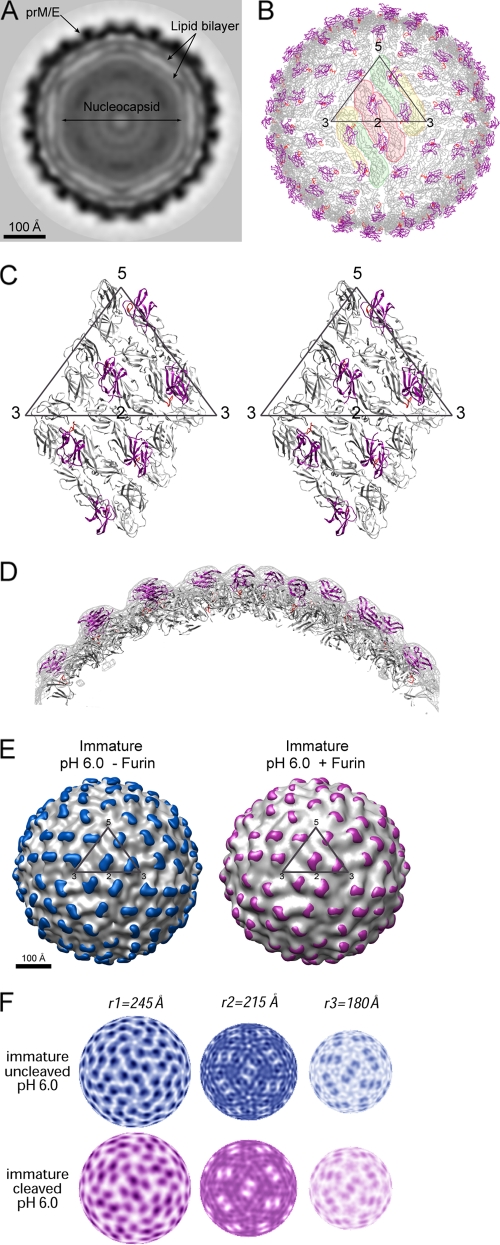
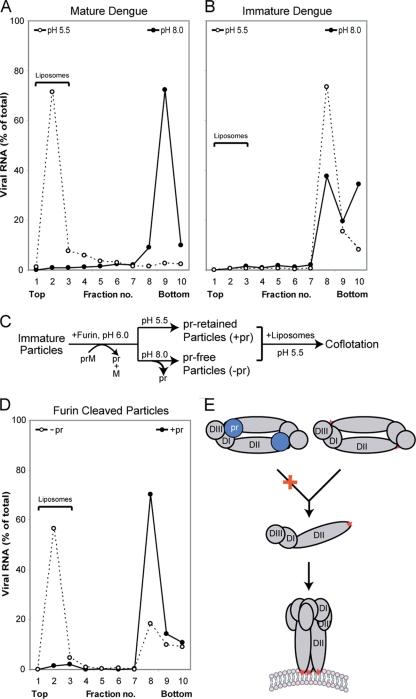
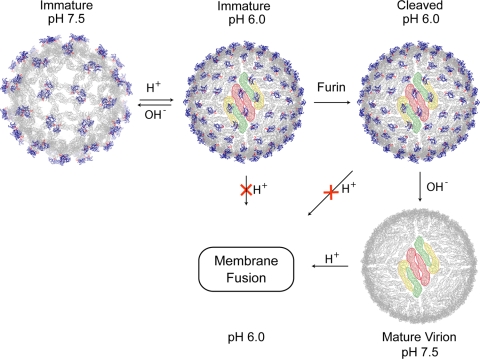
Flavivirus assembles into an inert particle that requires proteolytic activation by furin to enable transmission to other hosts. We previously showed that immature virus undergoes a conformational change at low pH that renders it accessible to furin (I. M. Yu, W. Zhang, H. A. Holdaway, L. Li, V. A. Kostyuchenko, P. R. Chipman, R. J. Kuhn, M. G. Rossmann, and J. Chen, Science 319:1834-1837, 2008). Here we show, using cryoelectron microscopy, that the structure of immature dengue virus at pH 6.0 is essentially the same before and after the cleavage of prM. The structure shows that after cleavage, the proteolytic product pr remains associated with the virion at acidic pH, and that furin cleavage by itself does not induce any major conformational changes. We also show by liposome cofloatation experiments that pr retention prevents membrane insertion, suggesting that pr is present on the virion in the trans-Golgi network to protect the progeny virus from fusion within the host cell.
Figures
References
-
- Baker, T. S., and R. H. Cheng. 1996. A model-based approach for determining orientations of biological macromolecules imaged by cryoelectron microscopy. J. Struct. Biol. 116:120-130. - PubMed
-
- Chen, J., K. H. Lee, D. A. Steinhauer, D. J. Stevens, J. J. Skehel, and D. C. Wiley. 1998. Structure of the hemagglutinin precursor cleavage site, a determinant of influenza pathogenicity and the origin of the labile conformation. Cell 95:409-417. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
