

Regulation of cell migration and morphogenesis by Abl-family kinases: emerging mechanisms and physiological contexts
- PMID: 19759284
- PMCID: PMC2746129
- DOI: 10.1242/jcs.039859
Regulation of cell migration and morphogenesis by Abl-family kinases: emerging mechanisms and physiological contexts
Abstract
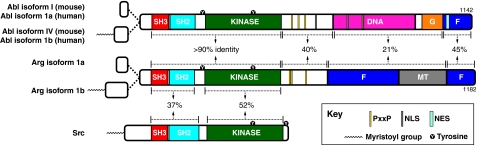
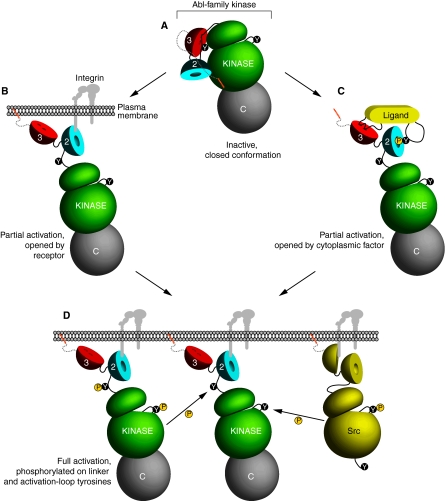
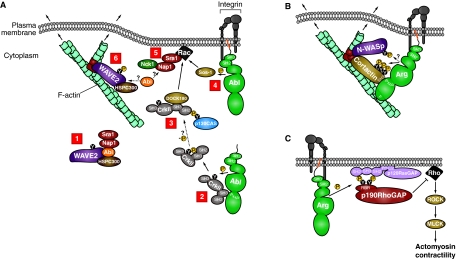
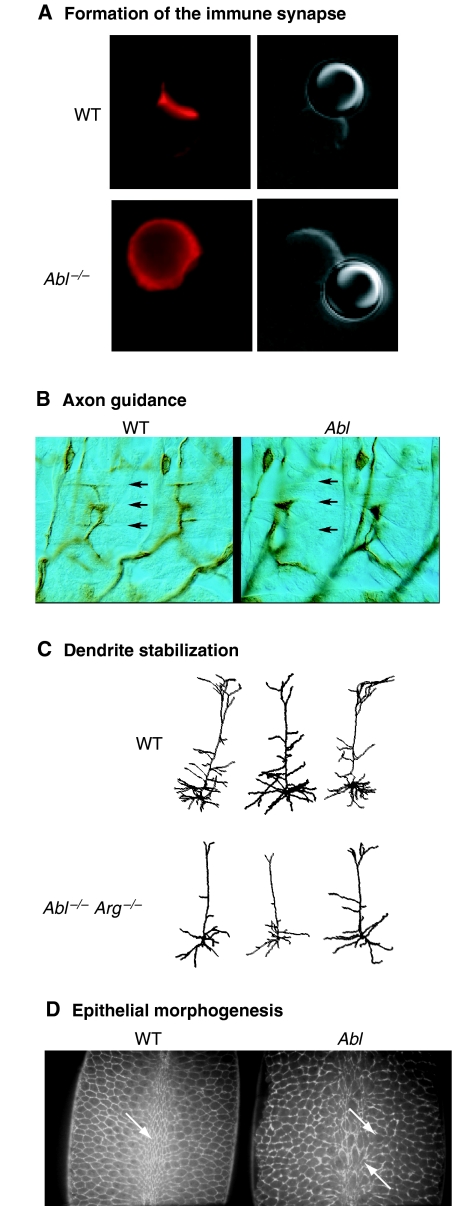
The Abl-family non-receptor tyrosine kinases are essential regulators of the cytoskeleton. They transduce diverse extracellular cues into cytoskeletal rearrangements that have dramatic effects on cell motility and morphogenesis. Recent biochemical and genetic studies have revealed several mechanisms that Abl-family kinases use to mediate these effects. Abl-family kinases stimulate actin polymerization through the activation of cortactin, hematopoietic lineage cell-specific protein (HS1), WASp- and WAVE-family proteins, and Rac1. They also attenuate cell contractility by inhibiting RhoA and altering adhesion dynamics. These pathways impinge on several physiological processes, including development and maintenance of the nervous and immune systems, and epithelial morphogenesis. Elucidating how Abl-family kinases are regulated, and where and when they coordinate cytoskeletal changes, is essential for garnering a better understanding of these complex processes.
Figures
Comment in
-
ABL at the leading edge.Nat Rev Mol Cell Biol. 2011 Jan;12(1):8. doi: 10.1038/nrm3026. Epub 2010 Nov 24. Nat Rev Mol Cell Biol. 2011. PMID: 21102623 No abstract available.
References
-
- Arthur, W. T., Noren, N. K. and Burridge, K. (2002). Regulation of Rho family GTPases by cell-cell and cell-matrix adhesion. Biol. Res. 35, 239-246. - PubMed
-
- Backert, S., Feller, S. M. and Wessler, S. (2008). Emerging roles of Abl family tyrosine kinases in microbial pathogenesis. Trends Biochem. Sci. 33, 80-90. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
