

Decisions in changing conditions: the urgency-gating model
- PMID: 19759303
- PMCID: PMC6665752
- DOI: 10.1523/JNEUROSCI.1844-09.2009
Decisions in changing conditions: the urgency-gating model
Abstract
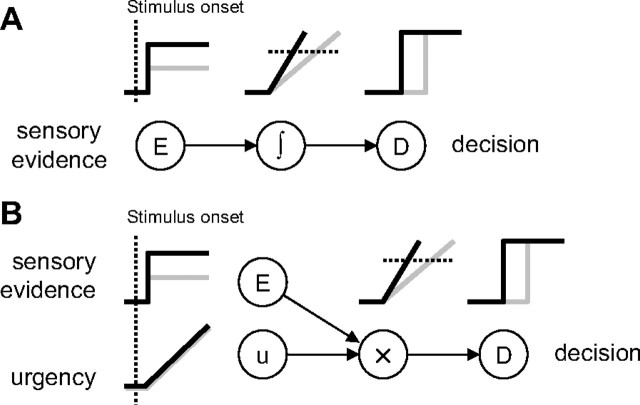
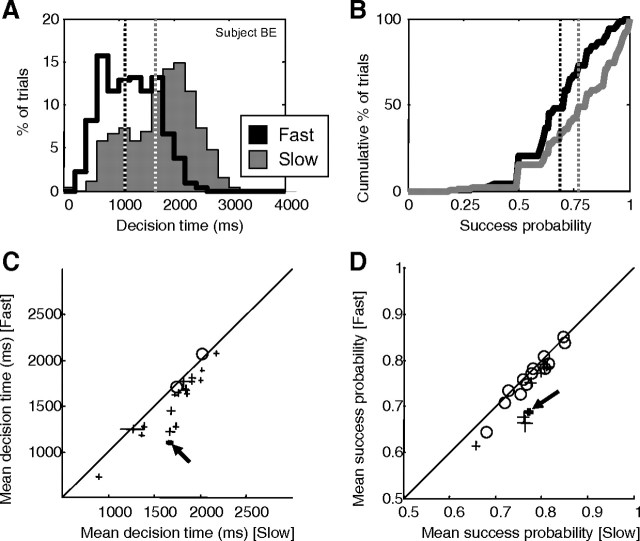
Several widely accepted models of decision making suggest that, during simple decision tasks, neural activity builds up until a threshold is reached and a decision is made. These models explain error rates and reaction time distributions in a variety of tasks and are supported by neurophysiological studies showing that neural activity in several cortical and subcortical regions gradually builds up at a rate related to task difficulty and reaches a relatively constant level of discharge at a time that predicts movement initiation. The mechanism responsible for this buildup is believed to be related to the temporal integration of sequential samples of sensory information. However, an alternative mechanism that may explain the neural and behavioral data is one in which the buildup of activity is instead attributable to a growing signal related to the urgency to respond, which multiplicatively modulates updated estimates of sensory evidence. These models are difficult to distinguish when, as in previous studies, subjects are presented with constant sensory evidence throughout each trial. To distinguish the models, we presented human subjects with a task in which evidence changed over the course of each trial. Our results are more consistent with "urgency gating" than with temporal integration of sensory samples and suggest a simple mechanism for implementing trade-offs between the speed and accuracy of decisions.
Figures







References
-
- Bogacz R, Gurney K. The basal ganglia and cortex implement optimal decision making between alternative actions. Neural Comput. 2007;19:442–477. - PubMed
-
- Bogacz R, Brown E, Moehlis J, Holmes P, Cohen JD. The physics of optimal decision making: a formal analysis of models of performance in two-alternative forced-choice tasks. Psychol Rev. 2006;113:700–765. - PubMed
-
- Carpenter RH, Williams ML. Neural computation of log likelihood in control of saccadic eye movements. Nature. 1995;377:59–62. - PubMed
-
- Chittka L, Skorupski P, Raine NE. Speed-accuracy tradeoffs in animal decision making. Trends Ecol Evol. 2009;24:400–407. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources