

Deletion-based reverse genetics in Medicago truncatula
- PMID: 19759346
- PMCID: PMC2773085
- DOI: 10.1104/pp.109.142919
Deletion-based reverse genetics in Medicago truncatula
Abstract
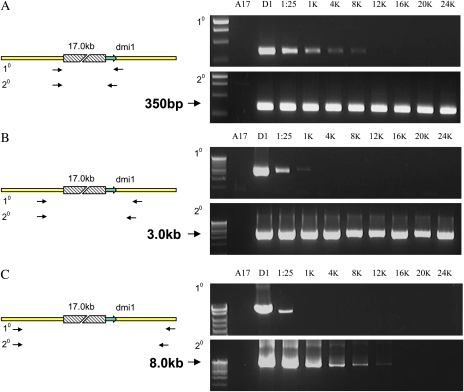
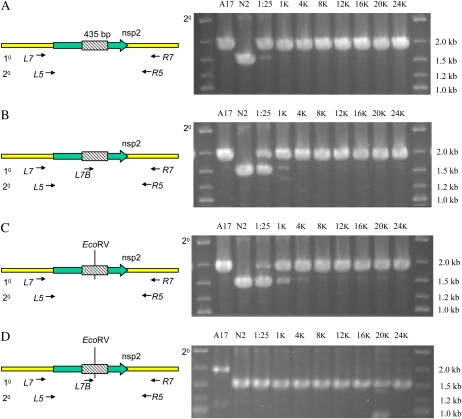
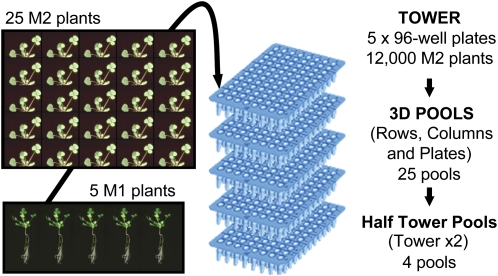
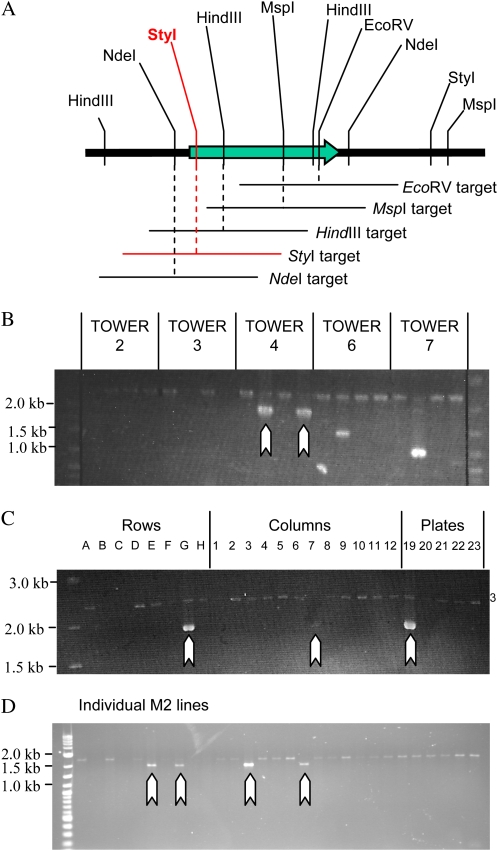
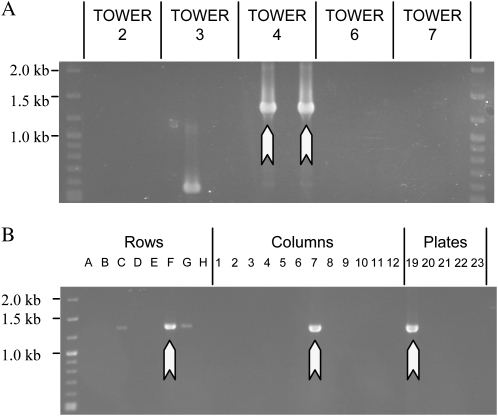
The primary goal of reverse genetics, the identification of null mutations in targeted genes, is achieved through screening large populations of randomly mutagenized plants. T-DNA and transposon-based mutagenesis has been widely employed but is limited to species in which transformation and tissue culture are efficient. In other species, TILLING (for Targeting Induced Local Lesions IN Genomes), based on chemical mutagenesis, has provided an efficient method for the identification of single base pair mutations, only 5% of which will be null mutations. Furthermore, the efficiency of inducing point mutations, like insertion-based mutations, is dependent on target size. Here, we describe an alternative reverse genetic strategy based on physically induced genomic deletions that, independent of target size, exclusively recovers knockout mutants. Deletion TILLING (De-TILLING) employs fast neutron mutagenesis and a sensitive polymerase chain reaction-based detection. A population of 156,000 Medicago truncatula plants has been structured as 13 towers each representing 12,000 M2 plants. The De-TILLING strategy allows a single tower to be screened using just four polymerase chain reaction reactions. Dual screening and three-dimensional pooling allows efficient location of mutants from within the towers. With this method, we have demonstrated the detection of mutants from this population at a rate of 29% using five targets per gene. This De-TILLING reverse genetic strategy is independent of tissue culture and efficient plant transformation and therefore applicable to any plant species. De-TILLING mutants offer advantages for crop improvement as they possess relatively few background mutations and no exogenous DNA.
Figures
Similar articles
-
TILLING: a shortcut in functional genomics.J Appl Genet. 2011 Nov;52(4):371-90. doi: 10.1007/s13353-011-0061-1. Epub 2011 Sep 13. J Appl Genet. 2011. PMID: 21912935 Free PMC article. Review.
-
Optimizing TILLING populations for reverse genetics in Medicago truncatula.Plant Biotechnol J. 2009 Jun;7(5):430-41. doi: 10.1111/j.1467-7652.2009.00410.x. Plant Biotechnol J. 2009. PMID: 19490506
-
Reverse genetics in Medicago truncatula using a TILLING mutant collection.Methods Mol Biol. 2013;1069:101-18. doi: 10.1007/978-1-62703-613-9_9. Methods Mol Biol. 2013. PMID: 23996312
-
Targeting Induced Local Lesions IN Genomes (TILLING) in Medicago truncatula.Methods Mol Biol. 2018;1822:71-82. doi: 10.1007/978-1-4939-8633-0_5. Methods Mol Biol. 2018. PMID: 30043297
-
Model Legumes: Functional Genomics Tools in Medicago truncatula.Methods Mol Biol. 2018;1822:11-37. doi: 10.1007/978-1-4939-8633-0_2. Methods Mol Biol. 2018. PMID: 30043294 Review.
Cited by
-
Keel petal incision: a simple and efficient method for genetic crossing in Medicago truncatula.Plant Methods. 2014 May 16;10:11. doi: 10.1186/1746-4811-10-11. eCollection 2014. Plant Methods. 2014. PMID: 24966878 Free PMC article.
-
Biological and genomic analysis of a symbiotic nitrogen fixation defective mutant in Medicago truncatula.Front Plant Sci. 2023 Jun 28;14:1209664. doi: 10.3389/fpls.2023.1209664. eCollection 2023. Front Plant Sci. 2023. PMID: 37457346 Free PMC article.
-
TILLING: a shortcut in functional genomics.J Appl Genet. 2011 Nov;52(4):371-90. doi: 10.1007/s13353-011-0061-1. Epub 2011 Sep 13. J Appl Genet. 2011. PMID: 21912935 Free PMC article. Review.
-
A draft genome of field pennycress (Thlaspi arvense) provides tools for the domestication of a new winter biofuel crop.DNA Res. 2015 Apr;22(2):121-31. doi: 10.1093/dnares/dsu045. Epub 2015 Jan 27. DNA Res. 2015. PMID: 25632110 Free PMC article.
-
Regulation of Compound Leaf Development.Plants (Basel). 2013 Dec 19;3(1):1-17. doi: 10.3390/plants3010001. Plants (Basel). 2013. PMID: 27135488 Free PMC article. Review.
References
-
- Alonso JM, Stepanova AN, Leisse TJ, Kim CJ, Chen HM, Shinn P, Stevenson DK, Zimmerman J, Barajas P, Cheuk R, et al (2003) Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 301 653–657 - PubMed
-
- Ane JM, Kiss GB, Riely BK, Penmetsa RV, Oldroyd GED, Ayax C, Levy J, Debelle F, Baek JM, Kalo P, et al (2004) Medicago truncatula DMI1 required for bacterial and fungal symbioses in legumes. Science 303 1364–1367 - PubMed
-
- Arabidopsis Genome Initiative (2000) Analysis of the genome sequence of the flowering plant Arabidopsis thaliana. Nature 408 796–815 - PubMed
-
- Azpiroz-Leehan R, Feldmann KA (1997) T-DNA insertion mutagenesis in Arabidopsis: going back and forth. Trends Genet 13 152–156 - PubMed
-
- Bruggemann E, Handwerger K, Essex C, Storz G (1996) Analysis of fast neutron-generated mutants at the Arabidopsis thaliana HY4 locus. Plant J 10 755–760 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
