

Stage-specific Arf tumor suppression in Notch1-induced T-cell acute lymphoblastic leukemia
- PMID: 19759355
- PMCID: PMC2777126
- DOI: 10.1182/blood-2009-07-233346
Stage-specific Arf tumor suppression in Notch1-induced T-cell acute lymphoblastic leukemia
Abstract
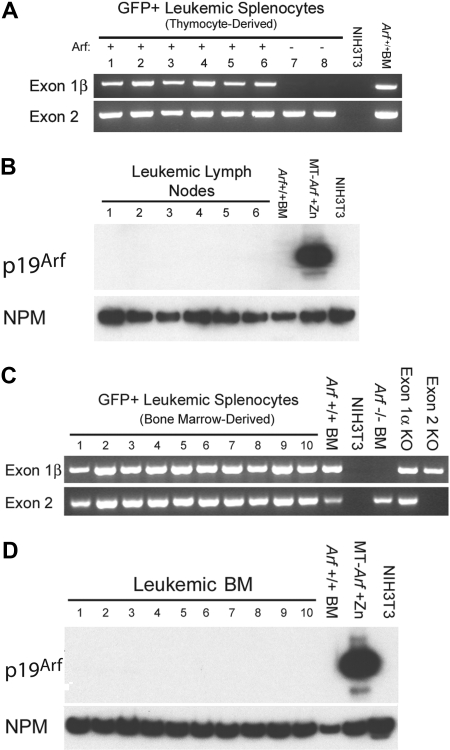
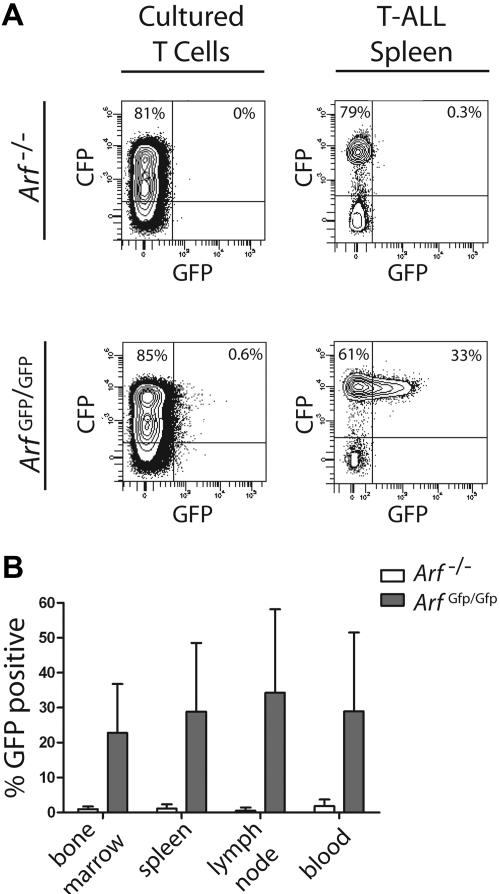
Frequent hallmarks of T-cell acute lymphoblastic leukemia (T-ALL) include aberrant NOTCH signaling and deletion of the CDKN2A locus, which contains 2 closely linked tumor suppressor genes (INK4A and ARF). When bone marrow cells or thymocytes transduced with a vector encoding the constitutively activated intracellular domain of Notch1 (ICN1) are expanded ex vivo under conditions that support T-cell development, cultured progenitors rapidly induce CD4+/CD8+ T-ALLs after infusion into healthy syngeneic mice. Under these conditions, enforced ICN1 expression also drives formation of T-ALLs in unconditioned CD-1 nude mice, bypassing any requirements for thymic maturation. Retention of Arf had relatively modest activity in suppressing the formation of T-ALLs arising from bone marrow-derived ICN1+ progenitors in which the locus is epigenetically silenced, and all resulting Arf (+/+) tumors failed to express the p19(Arf) protein. In striking contrast, retention of Arf in thymocyte-derived ICN1+ donor cells significantly delayed disease onset and suppressed the penetrance of T-ALL. Use of cultured thymocyte-derived donor cells expressing a functionally null Arf-GFP knock-in allele confirmed that ICN1 signaling can induce Arf expression in vivo. Arf activation by ICN1 in T cells thereby provides stage-specific tumor suppression but also a strong selective pressure for deletion of the locus in T-ALL.
Figures
References
-
- Pui C-H, Robison LL, Look AT. Acute lymphoblastic leukemia. Lancet. 2008;371(9617):1030–1043. - PubMed
-
- Weng AP, Ferrando AA, Lee W, et al. Activating mutations of NOTCH1 in human T cell acute lymphoblastic leukemia. Science. 2004;306(5694):269–271. - PubMed
-
- Irvine KD, Rauskolb C. Boundaries in development: formation and function. Annu Rev Cell Dev Biol. 2001;17:189–214. - PubMed
-
- Grabher C, von Boehmer H, Look AT. Notch 1 activation in the molecular pathogenesis of T-cell acute lymphoblastic leukemia. Nat Rev Cancer. 2006;6(5):347–359. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
