

Genomic analysis suggests that mRNA destabilization by the microprocessor is specialized for the auto-regulation of Dgcr8
- PMID: 19759829
- PMCID: PMC2736397
- DOI: 10.1371/journal.pone.0006971
Genomic analysis suggests that mRNA destabilization by the microprocessor is specialized for the auto-regulation of Dgcr8
Abstract
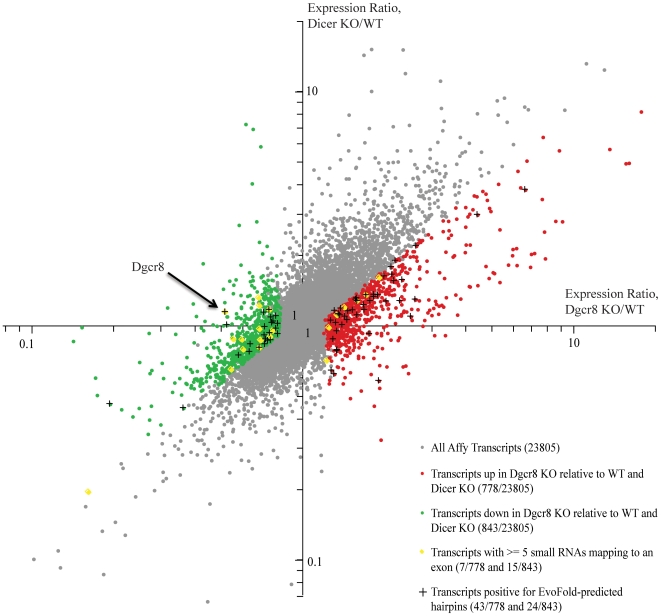
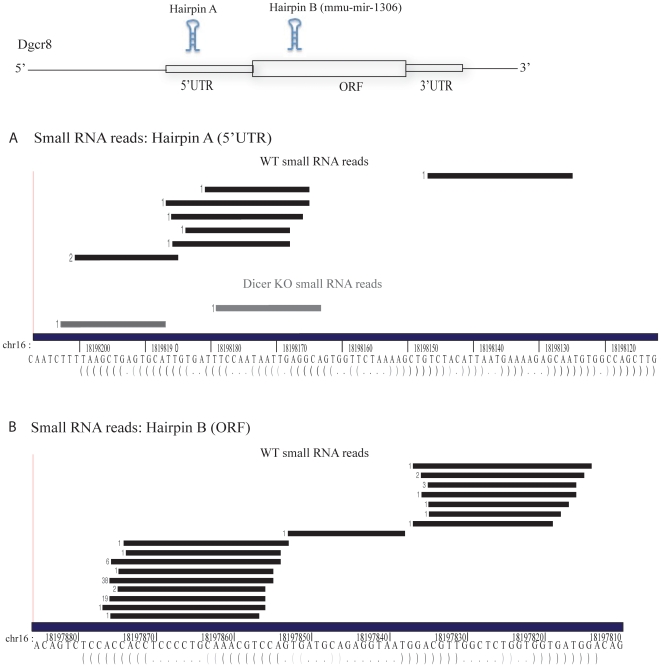
Background: The Microprocessor, containing the RNA binding protein Dgcr8 and RNase III enzyme Drosha, is responsible for processing primary microRNAs to precursor microRNAs. The Microprocessor regulates its own levels by cleaving hairpins in the 5'UTR and coding region of the Dgcr8 mRNA, thereby destabilizing the mature transcript.
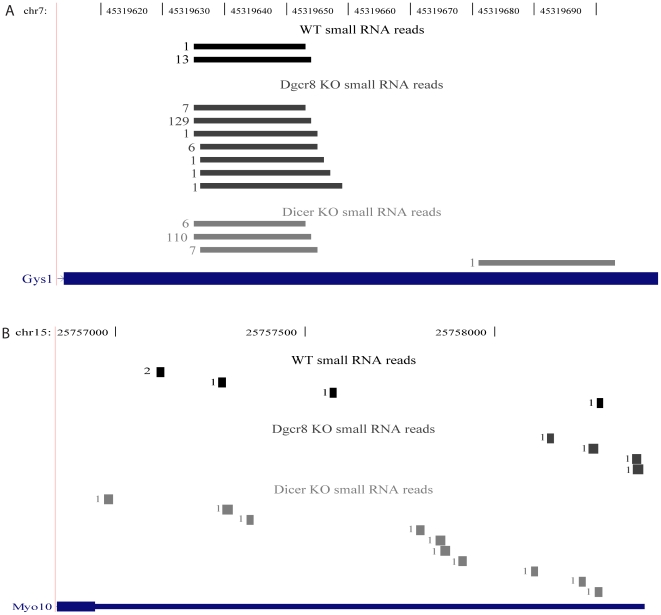
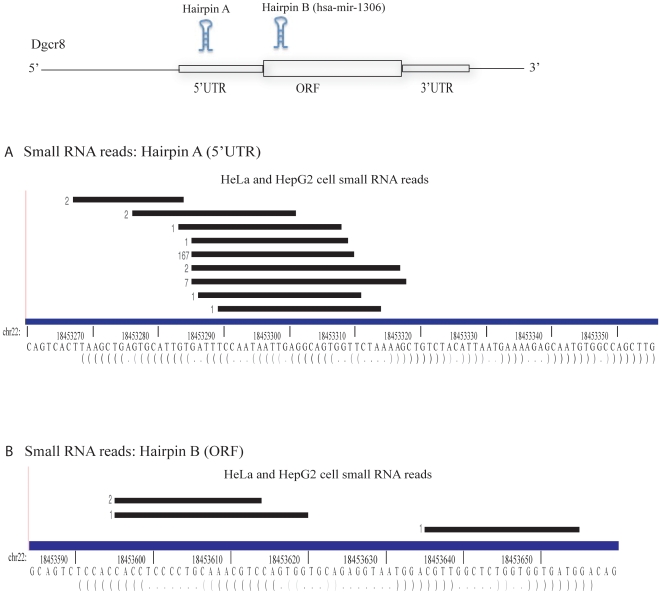
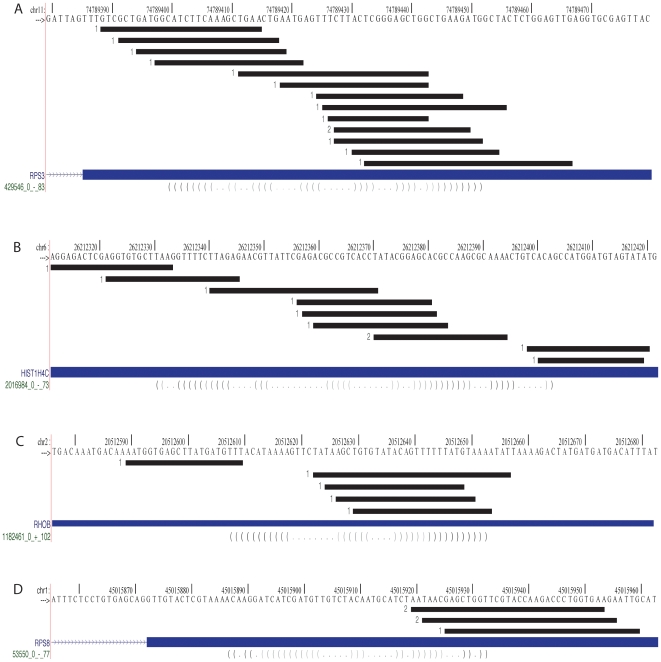
Methodology/principal findings: To determine whether the Microprocessor has a broader role in directly regulating other coding mRNA levels, we integrated results from expression profiling and ultra high-throughput deep sequencing of small RNAs. Expression analysis of mRNAs in wild-type, Dgcr8 knockout, and Dicer knockout mouse embryonic stem (ES) cells uncovered mRNAs that were specifically upregulated in the Dgcr8 null background. A number of these transcripts had evolutionarily conserved predicted hairpin targets for the Microprocessor. However, analysis of deep sequencing data of 18 to 200nt small RNAs in mouse ES, HeLa, and HepG2 indicates that exonic sequence reads that map in a pattern consistent with Microprocessor activity are unique to Dgcr8.
Conclusion/significance: We conclude that the Microprocessor's role in directly destabilizing coding mRNAs is likely specifically targeted to Dgcr8 itself, suggesting a specialized cellular mechanism for gene auto-regulation.
Conflict of interest statement
Figures
References
-
- Babiarz JE, Blelloch R. Small RNAs – their biogenesis, regulation and function in embryonic stem cells. 2009. In StemBook ( www.stembook.org/node/583) - PubMed
-
- Lee Y, Ahn C, Han J, Choi H, Kim J, et al. The nuclear RNase III Drosha initiates microRNA processing. Nature. 2003;425:415–419. - PubMed
-
- Denli AM, Tops BB, Plasterk RH, Ketting RF, Hannon GJ. Processing of primary microRNAs by the Microprocessor complex. Nature. 2004;432:231–235. - PubMed
-
- Gregory RI, Yan KP, Amuthan G, Chendrimada T, Doratotaj B, et al. The Microprocessor complex mediates the genesis of microRNAs. Nature. 2004;432:235–240. - PubMed
