

Mitochondrial localization of PARP-1 requires interaction with mitofilin and is involved in the maintenance of mitochondrial DNA integrity
- PMID: 19762472
- PMCID: PMC2797232
- DOI: 10.1074/jbc.M109.025882
Mitochondrial localization of PARP-1 requires interaction with mitofilin and is involved in the maintenance of mitochondrial DNA integrity
Abstract
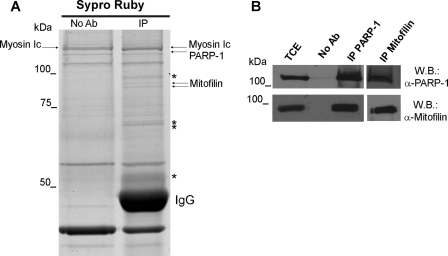
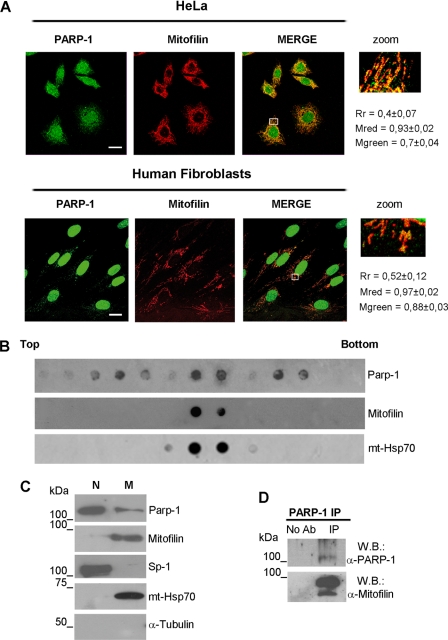
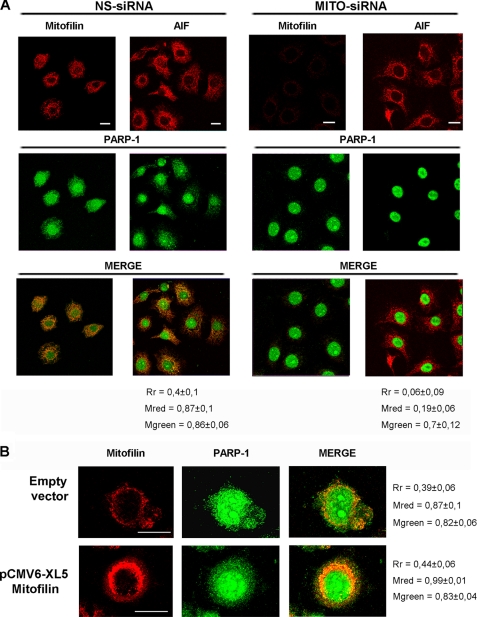
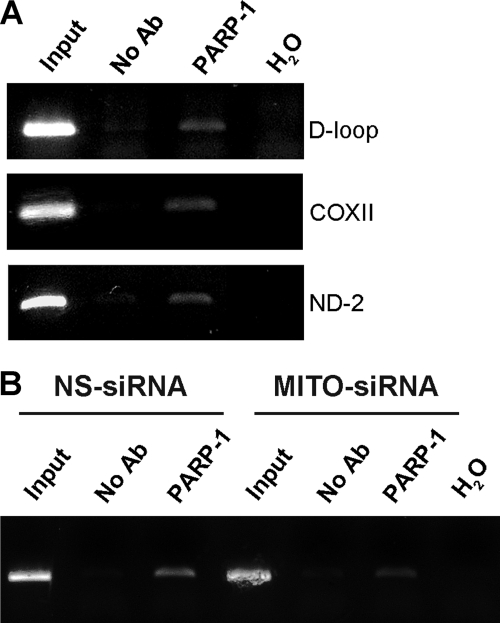
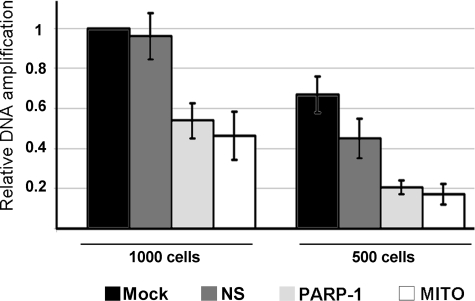
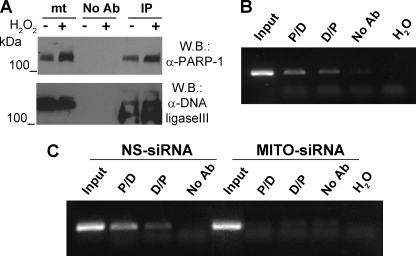
Poly(ADP-ribose)polymerase-1 (PARP-1) is a predominantly nuclear enzyme that exerts numerous functions in cellular physiology and pathology, from maintenance of DNA stability to transcriptional regulation. Through a proteomic analysis of PARP-1 co-immunoprecipitation complexes, we identified Mitofilin, a mitochondrial protein, as a new PARP-1 interactor. This result prompted us to further investigate the presence and the role of the enzyme in mitochondria. Using laser confocal microscopy and Western blot analysis of purified mitochondria, we demonstrated the mitochondrial localization of a fraction of PARP-1. Further, the effects of overexpressing or down-regulating Mitofilin showed that this protein promotes and is required for PARP-1 mitochondrial localization. We also report several lines of evidence suggesting that intramitochondrial PARP-1 plays a role in mitochondrial DNA (mtDNA) damage signaling and/or repair. First, we show that PARP-1 binds to different regions throughout the mtDNA. Moreover, we demonstrated that the depletion of either PARP-1 or Mitofilin, which abrogates the mitochondrial localization of the enzyme, leads to the accumulation of mtDNA damage. Finally, we show that DNA ligase III, known to be required for mtDNA repair, participates in a PARP-1-containing complex bound to mtDNA. This work highlights a new environment for PARP-1, opening the possibility that at least some of the nuclear functions of the enzyme can be also extended to mtDNA metabolism.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
