

Ligand-regulated oligomerization of beta(2)-adrenoceptors in a model lipid bilayer
- PMID: 19763081
- PMCID: PMC2748299
- DOI: 10.1038/emboj.2009.267
Ligand-regulated oligomerization of beta(2)-adrenoceptors in a model lipid bilayer
Abstract
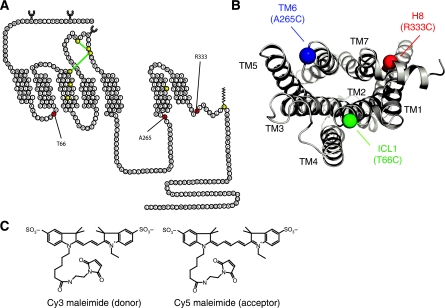
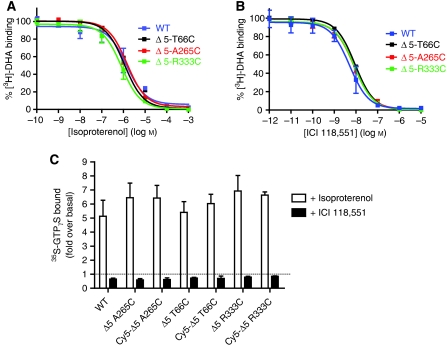
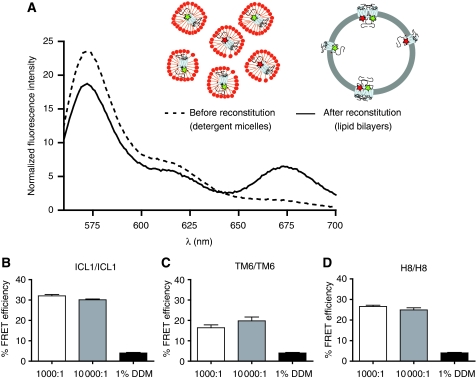
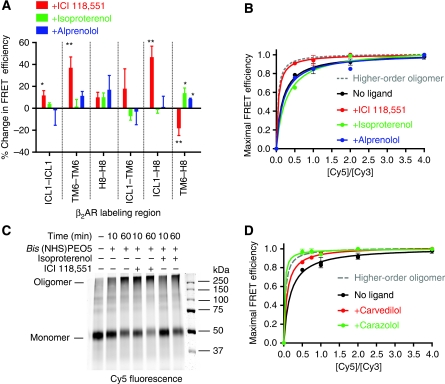
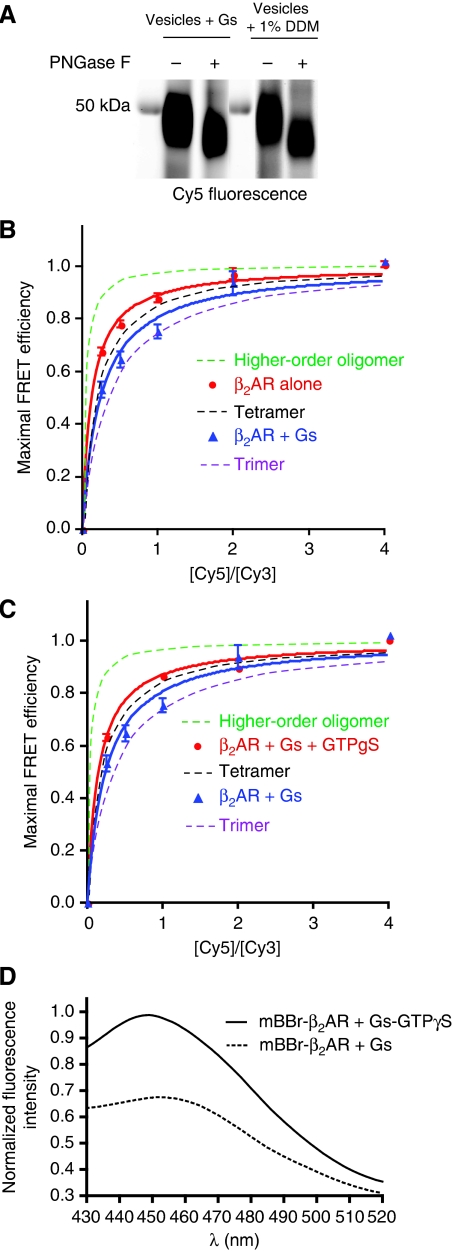
The beta(2)-adrenoceptor (beta(2)AR) was one of the first Family A G protein-coupled receptors (GPCRs) shown to form oligomers in cellular membranes, yet we still know little about the number and arrangement of protomers in oligomers, the influence of ligands on the organization or stability of oligomers, or the requirement for other proteins to promote oligomerization. We used fluorescence resonance energy transfer (FRET) to characterize the oligomerization of purified beta(2)AR site-specifically labelled at three different positions with fluorophores and reconstituted into a model lipid bilayer. Our results suggest that the beta(2)AR is predominantly tetrameric following reconstitution into phospholipid vesicles. Agonists and antagonists have little effect on the relative orientation of protomers in oligomeric complexes. In contrast, binding of inverse agonists leads to significant increases in FRET efficiencies for most labelling pairs, suggesting that this class of ligand promotes tighter packing of protomers and/or the formation of more complex oligomers by reducing conformational fluctuations in individual protomers. The results provide new structural insights into beta(2)AR oligomerization and suggest a possible mechanism for the functional effects of inverse agonists.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures




References
-
- Allen JA, Halverson-Tamboli RA, Rasenick MM (2007) Lipid raft microdomains and neurotransmitter signalling. Nat Rev Neurosci 8: 128–140 - PubMed
-
- Angers S, Bouvier M (2000) Reply: beyond receptor dimerization. Trends Pharmacol Sci 21: 326. - PubMed
-
- Bayburt TH, Leitz AJ, Xie G, Oprian DD, Sligar SG (2007) Transducin activation by nanoscale lipid bilayers containing one and two rhodopsins. J Biol Chem 282: 14875–14881 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
