

NEMO specifically recognizes K63-linked poly-ubiquitin chains through a new bipartite ubiquitin-binding domain
- PMID: 19763089
- PMCID: PMC2760117
- DOI: 10.1038/emboj.2009.241
NEMO specifically recognizes K63-linked poly-ubiquitin chains through a new bipartite ubiquitin-binding domain
Abstract
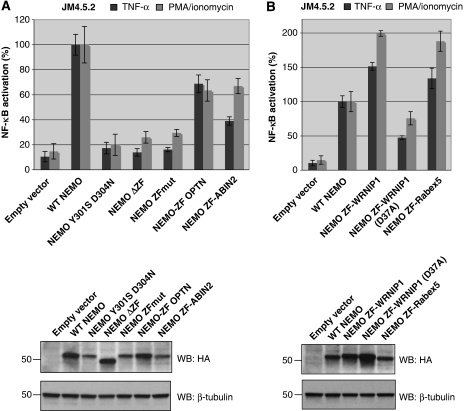
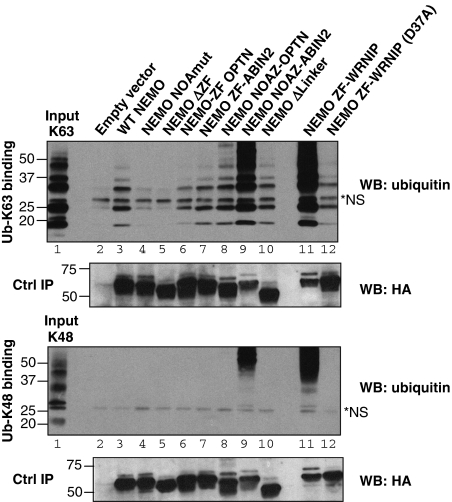
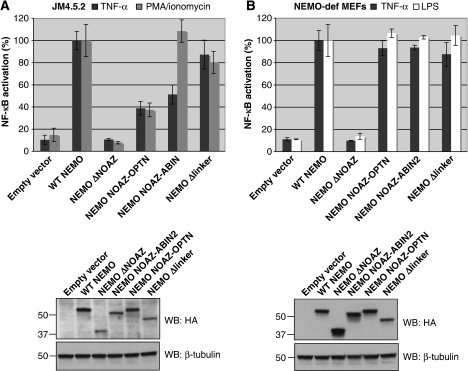
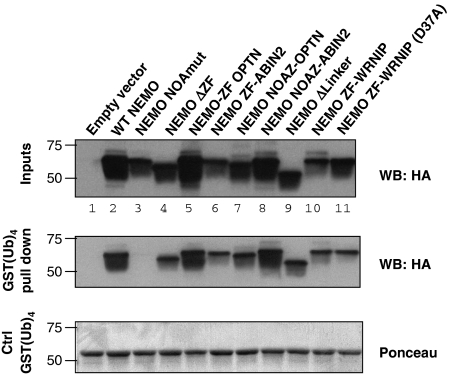
An important property of NEMO, the core element of the IKK complex involved in NF-kappaB activation, resides in its ability to specifically recognize poly-ubiquitin chains. A small domain called NOA/UBAN has been suggested to be responsible for this property. We recently demonstrated that the C-terminal Zinc Finger (ZF) of NEMO is also able to bind ubiquitin. We show here by ZF swapping and mutagenesis that this represents its only function. While neither NOA nor ZF shows any preference for K63-linked chains, we demonstrate that together they form a bipartite high-affinity K63-specific ubiquitin-binding domain. A similar domain can be found in two other proteins, Optineurin and ABIN2, and can be freely exchanged with that of NEMO without interfering with its activity. This suggests that the main function of the C-terminal half of NEMO is to specifically bind K63-linked poly-ubiquitin chains. We also demonstrate that the recently described binding of NEMO to linear poly-ubiquitin chains is dependent on the NOA alone and does not require the presence of the ZF.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures

Similar articles
-
Two-sided ubiquitin binding of NF-κB essential modulator (NEMO) zinc finger unveiled by a mutation associated with anhidrotic ectodermal dysplasia with immunodeficiency syndrome.J Biol Chem. 2013 Nov 22;288(47):33722-33737. doi: 10.1074/jbc.M113.483305. Epub 2013 Oct 7. J Biol Chem. 2013. PMID: 24100029 Free PMC article.
-
Covalent modification of the NF-κB essential modulator (NEMO) by a chemical compound can regulate its ubiquitin binding properties in vitro.J Biol Chem. 2014 Nov 28;289(48):33161-74. doi: 10.1074/jbc.M114.582478. Epub 2014 Oct 8. J Biol Chem. 2014. PMID: 25296760 Free PMC article.
-
Direct inhibition of NF-κB activation by peptide targeting the NOA ubiquitin binding domain of NEMO.Biochem Pharmacol. 2011 Nov 1;82(9):1163-74. doi: 10.1016/j.bcp.2011.07.083. Epub 2011 Jul 23. Biochem Pharmacol. 2011. PMID: 21803029
-
NEMO Family of Proteins as Polyubiquitin Receptors: Illustrating Non-Degradative Polyubiquitination's Roles in Health and Disease.Cells. 2025 Feb 18;14(4):304. doi: 10.3390/cells14040304. Cells. 2025. PMID: 39996775 Free PMC article. Review.
-
Linear polyubiquitination: a new regulator of NF-kappaB activation.EMBO Rep. 2009 Jul;10(7):706-13. doi: 10.1038/embor.2009.144. Epub 2009 Jun 19. EMBO Rep. 2009. PMID: 19543231 Free PMC article. Review.
Cited by
-
Impairment of protein trafficking upon overexpression and mutation of optineurin.PLoS One. 2010 Jul 12;5(7):e11547. doi: 10.1371/journal.pone.0011547. PLoS One. 2010. PMID: 20634958 Free PMC article.
-
The Many Roles of Ubiquitin in NF-κB Signaling.Biomedicines. 2018 Apr 10;6(2):43. doi: 10.3390/biomedicines6020043. Biomedicines. 2018. PMID: 29642643 Free PMC article. Review.
-
NF-κB: At the Borders of Autoimmunity and Inflammation.Front Immunol. 2021 Aug 9;12:716469. doi: 10.3389/fimmu.2021.716469. eCollection 2021. Front Immunol. 2021. PMID: 34434197 Free PMC article. Review.
-
A ubiquitin replacement strategy in human cells reveals distinct mechanisms of IKK activation by TNFalpha and IL-1beta.Mol Cell. 2009 Oct 23;36(2):302-14. doi: 10.1016/j.molcel.2009.10.002. Mol Cell. 2009. PMID: 19854138 Free PMC article.
-
Linear ubiquitination prevents inflammation and regulates immune signalling.Nature. 2011 Mar 31;471(7340):591-6. doi: 10.1038/nature09816. Nature. 2011. PMID: 21455173
References
-
- Bagnéris C, Ageichik AV, Cronin N, Wallace B, Collins M, Boshoff C, Waksman G, Barrett T (2008) Crystal structure of a vFlip–IKKgamma complex: insights into viral activation of the IKK signalosome. Mol Cell 30: 620–631 - PubMed
-
- Bienko M, Green CM, Crosetto N, Rudolf F, Zapart G, Coull B, Kannouche P, Wider G, Peter M, Lehmann AR, Hofmann K, Dikic I (2005) Ubiquitin-binding domains in Y-family polymerases regulate translesion synthesis. Science 310: 1821–1824 - PubMed
-
- Bish RA, Fregoso OI, Piccini A, Myers MP (2008) Conjugation of complex polyubiquitin chains to WRNIP1. J Proteome Res 7: 3481–3489 - PubMed
-
- Bish RA, Myers MP (2007) Werner helicase-interacting protein 1 binds polyubiquitin via its zinc finger domain. J Biol Chem 282: 23184–23193 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
