

Broad-scale recombination patterns underlying proper disjunction in humans
- PMID: 19763175
- PMCID: PMC2734982
- DOI: 10.1371/journal.pgen.1000658
Broad-scale recombination patterns underlying proper disjunction in humans
Abstract
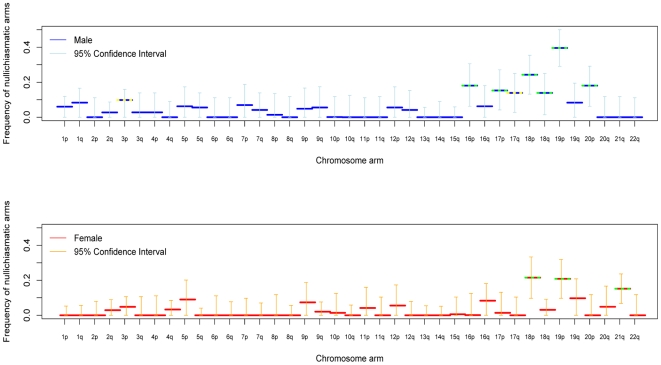
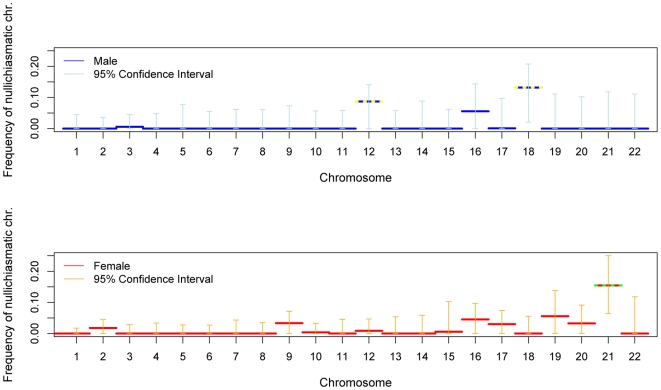
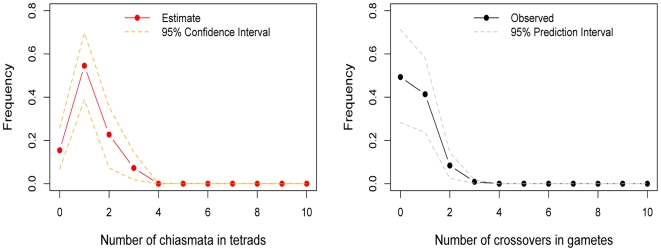
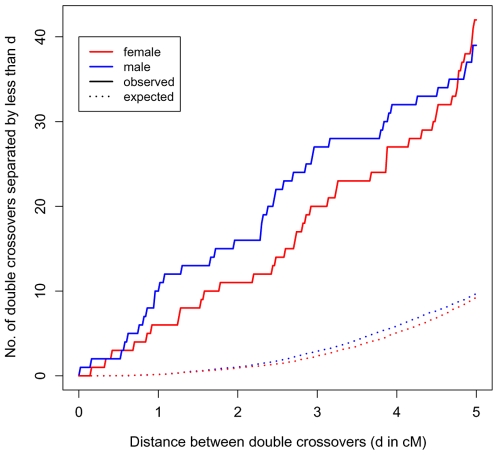
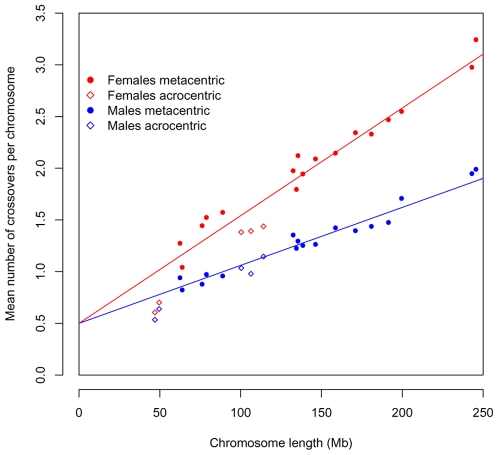
Although recombination is essential to the successful completion of human meiosis, it remains unclear how tightly the process is regulated and over what scale. To assess the nature and stringency of constraints on human recombination, we examined crossover patterns in transmissions to viable, non-trisomic offspring, using dense genotyping data collected in a large set of pedigrees. Our analysis supports a requirement for one chiasma per chromosome rather than per arm to ensure proper disjunction, with additional chiasmata occurring in proportion to physical length. The requirement is not absolute, however, as chromosome 21 seems to be frequently transmitted properly in the absence of a chiasma in females, a finding that raises the possibility of a back-up mechanism aiding in its correct segregation. We also found a set of double crossovers in surprisingly close proximity, as expected from a second pathway that is not subject to crossover interference. These findings point to multiple mechanisms that shape the distribution of crossovers, influencing proper disjunction in humans.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Otto SP, Lenormand T. Resolving the paradox of sex and recombination. Nat Rev Genet. 2002;3:252–261. - PubMed
-
- Hunt PA, Hassold TJ. Sex matters in meiosis. Science. 2002;296:2181–2183. - PubMed
-
- Hassold T, Hunt P. To err (meiotically) is human: the genesis of human aneuploidy. Nat Rev Genet. 2001;2:280–291. - PubMed
-
- Kong A, Barnard J, Gudbjartsson DF, Thorleifsson G, Jonsdottir G, et al. Recombination rate and reproductive success in humans. Nat Genet. 2004;36:1203–1206. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
