

Anti-CD44 induces apoptosis in T lymphoma via mitochondrial depolarization
- PMID: 19765170
- PMCID: PMC3829012
- DOI: 10.1111/j.1582-4934.2009.00909.x
Anti-CD44 induces apoptosis in T lymphoma via mitochondrial depolarization
Abstract
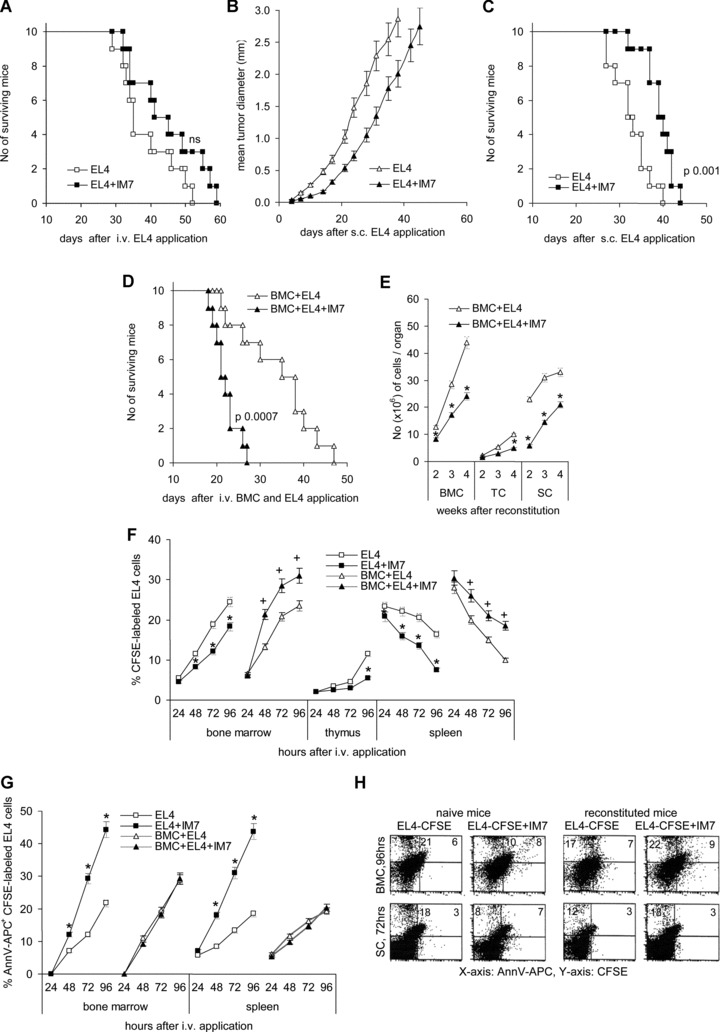
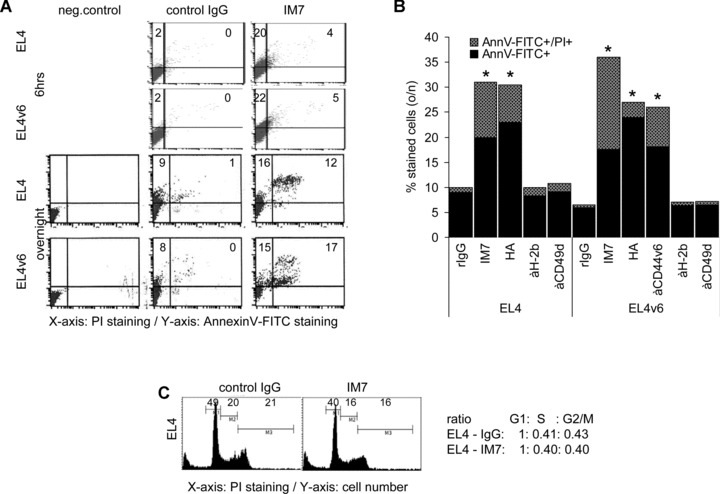
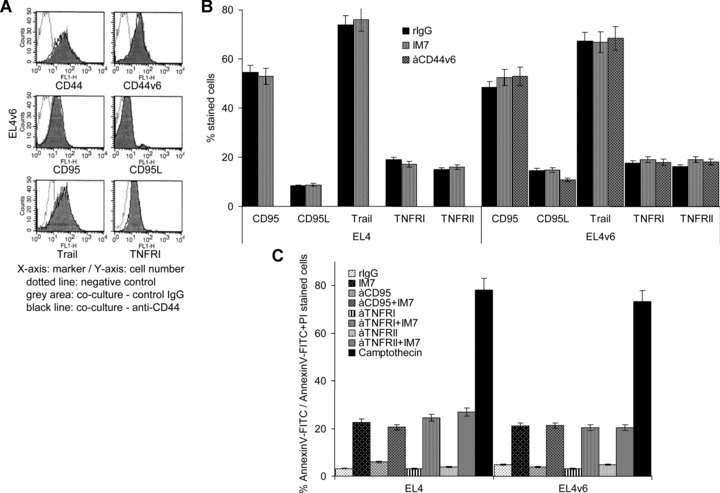
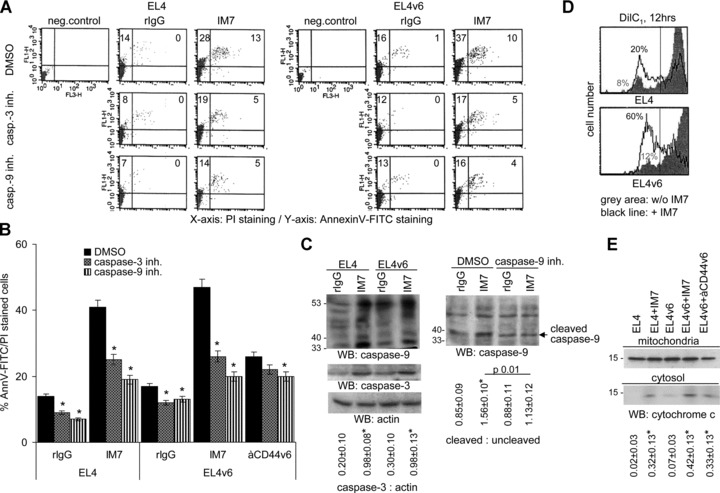
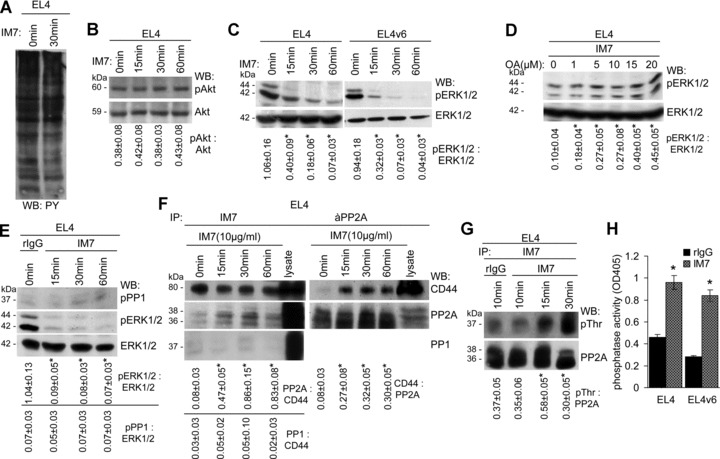
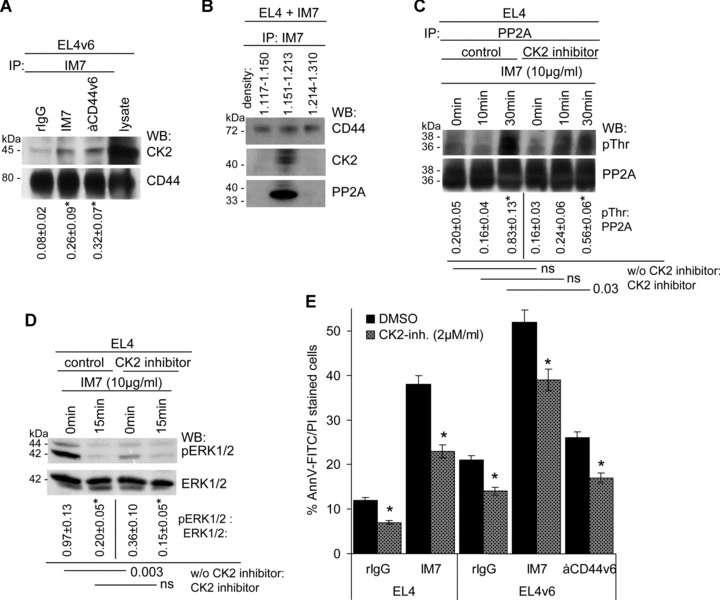
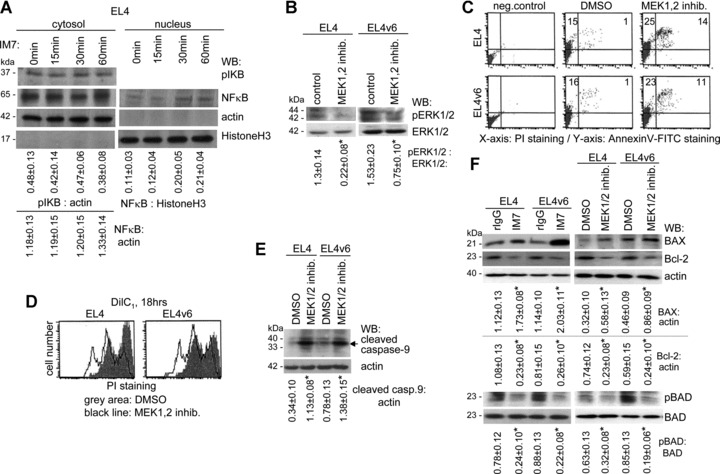
A blockade of CD44 can interfere with haematopoietic and leukemic stem cell homing, the latter being considered as a therapeutic option in haematological malignancies. We here aimed to explore the molecular mechanism underlying the therapeutic efficacy of anti-CD44. We noted that in irradiated mice reconstituted with a bone marrow cell transplant, anti-CD44 exerts a stronger effect on haematopoietic reconstitution than on T lymphoma (EL4) growth. Nonetheless, in the non-reconstituted mouse anti-CD44 suffices for a prolonged survival of EL4-bearing mice, where anti-CD44-prohibited homing actively drives EL4 cells into apoptosis. In vitro, a CD44 occupancy results in a 2-4-fold increase in apoptotic EL4 cells. Death receptor expression (CD95, TRAIL, TNFRI) remains unaltered and CD95 cross-linking-mediated apoptosis is not affected. Instead, CD44 ligation promotes mitochondrial depolarization that is accompanied by caspase-9 cleavage and is inhibited in the presence of a caspase-9 inhibitor. Apoptosis becomes initiated by activation of CD44-associated phosphatase 2A (PP2A) and proceeds via ERK1/2 dephosphorylation without ERK1/2 degradation. Accordingly, CD44-induced apoptosis could be mimicked by ERK1/2 inhibition, that also promotes EL4 cell apoptosis through the mitochondrial pathway. Thus, during haematopoietic stem cell reconstitution care should be taken not to interfere by a blockade of CD44 with haematopoiesis, which could be circumvented by selectively targeting leukemic CD44 isoforms. Beyond homing/settlement in the bone marrow niche, anti-CD44 drives leukemic T cells into apoptosis via the mitochondrial death pathway by CD44 associating with PP2A. Uncovering this new pathway of CD44-induced leukemic cell death provides new options of therapeutic interference.
Figures
References
-
- Lynch KW. Consequences of regulated pre-mRNA splicing in the immune system. Nat Rev Immunol. 2004;4:931–40. - PubMed
-
- Naor D, Wallach-Dayan SB, Zahalka MA, Sionov RV. Involvement of CD44, a molecule with a thousand faces, in cancer dissemination. Semin Cancer Biol. 2008;18:260–7. - PubMed
-
- Orian-Rousseau V, Ponta H. Adhesion proteins meet receptors: a common theme. Adv Cancer Res. 2008;101:63–92. - PubMed
-
- Lapidot T, Dar A, Kollet O. How do stem cells find their way home? Blood. 2005;106:1901–10. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
