

A dual role for the S-locus receptor kinase in self-incompatibility and pistil development revealed by an Arabidopsis rdr6 mutation
- PMID: 19767457
- PMCID: PMC2768922
- DOI: 10.1105/tpc.109.067801
A dual role for the S-locus receptor kinase in self-incompatibility and pistil development revealed by an Arabidopsis rdr6 mutation
Abstract
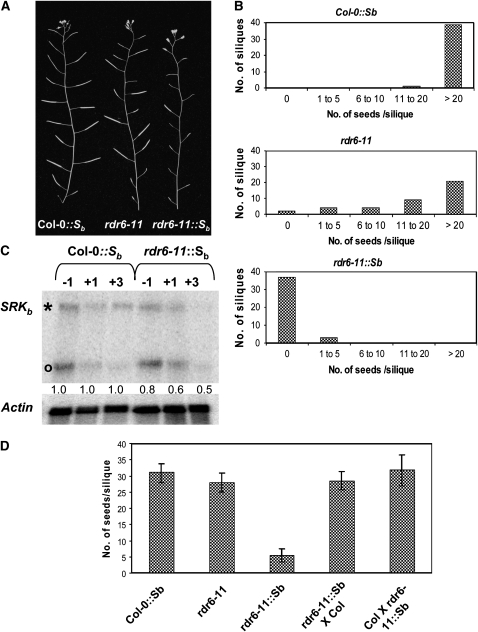
The coordinate evolution of self-incompatibility (SI) and stigma-anther separation, two mechanisms that promote cross-pollination in plants, has been a long-standing puzzle in evolution and development. Using a transgenic self-incompatible Arabidopsis thaliana model, we performed screens for mutants exhibiting a modified SI response. A mutation in the RNA-dependent RNA polymerase RDR6, which functions in trans-acting short interfering RNA (ta-siRNA) production, was found that simultaneously enhances SI and causes stigma exsertion, without associated increases in SRK transcript levels. While rdr6 mutants had been previously shown to exhibit stochastic stigma exsertion, our results demonstrate that the S-locus receptor kinase (SRK) gene further enhances pistil elongation and stigma exsertion in this mutant background, a process that requires SRK catalytic activity and correlates with SRK transcript levels. These results suggest that positive regulators or effectors of SI and pistil development are regulated by ta-siRNA(s). By establishing complex connections between SI and stigma exsertion through the sharing of a ta-siRNA-mediated regulatory pathway and the dual role of SRK in SI and pistil development, our study provides a molecular explanation for the coordinate evolution of these processes.
Figures







References
-
- Adenot, X., Elmayan, T., Lauressergues, D., Boutet, S., Bouche, N., Gasciolli, V., and Vaucheret, H. (2006). DRB4-dependent TAS3 trans-acting siRNAs control leaf morphology through AGO7. Curr. Biol. 16 927–932. - PubMed
-
- Arias, A.M., and Hayward, P. (2006). Filtering transcriptional noise during development: concepts and mechanisms. Nat. Rev. Genet. 7 34–44. - PubMed
-
- Axtell, M.J., and Bowman, J.L. (2008). Evolution of plant microRNAs and their targets. Trends Plant Sci. 13 343–349. - PubMed
-
- Barrett, S.C. (2002). The evolution of plant sexual diversity. Nat. Rev. Genet. 3 274–284. - PubMed
-
- Bjorkman, T. (1995). The effectiveness of heterostyly in preventing illegitimate pollination in dish-shaped flowers. Sex. Plant Reprod. 8 143–149.
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
