

Structural and kinetic determinants of protease substrates
- PMID: 19767749
- PMCID: PMC4042863
- DOI: 10.1038/nsmb.1668
Structural and kinetic determinants of protease substrates
Abstract
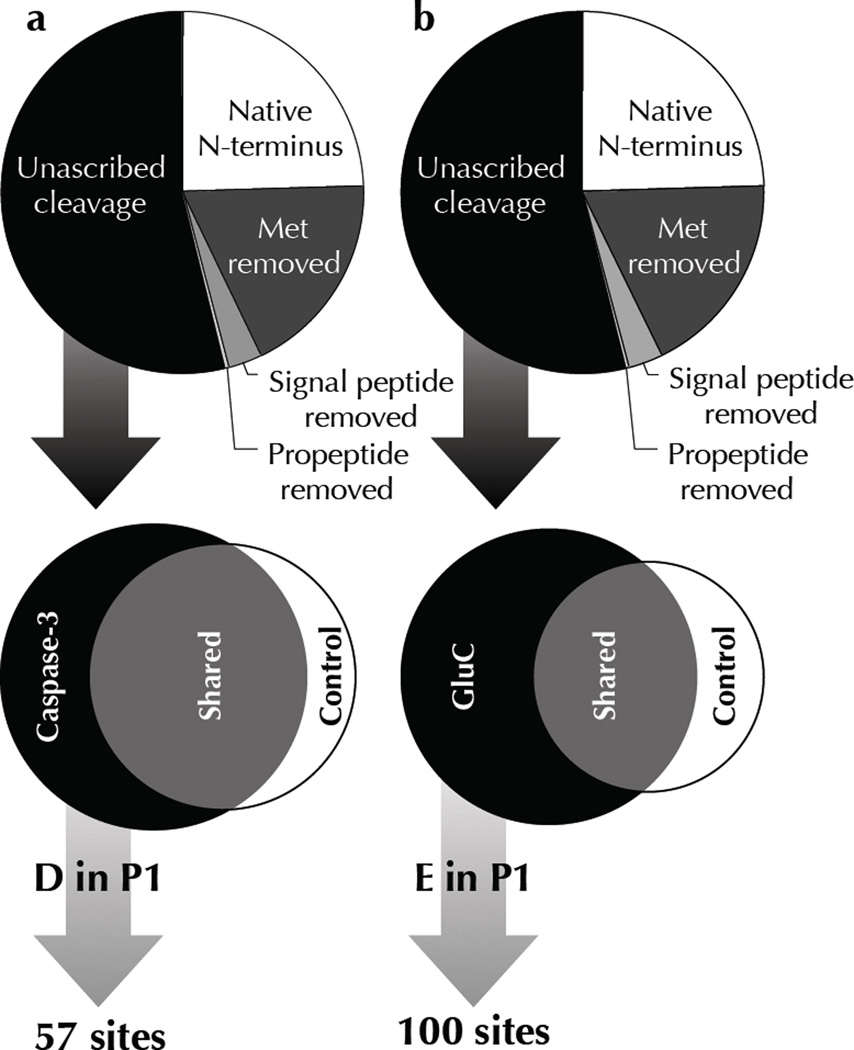
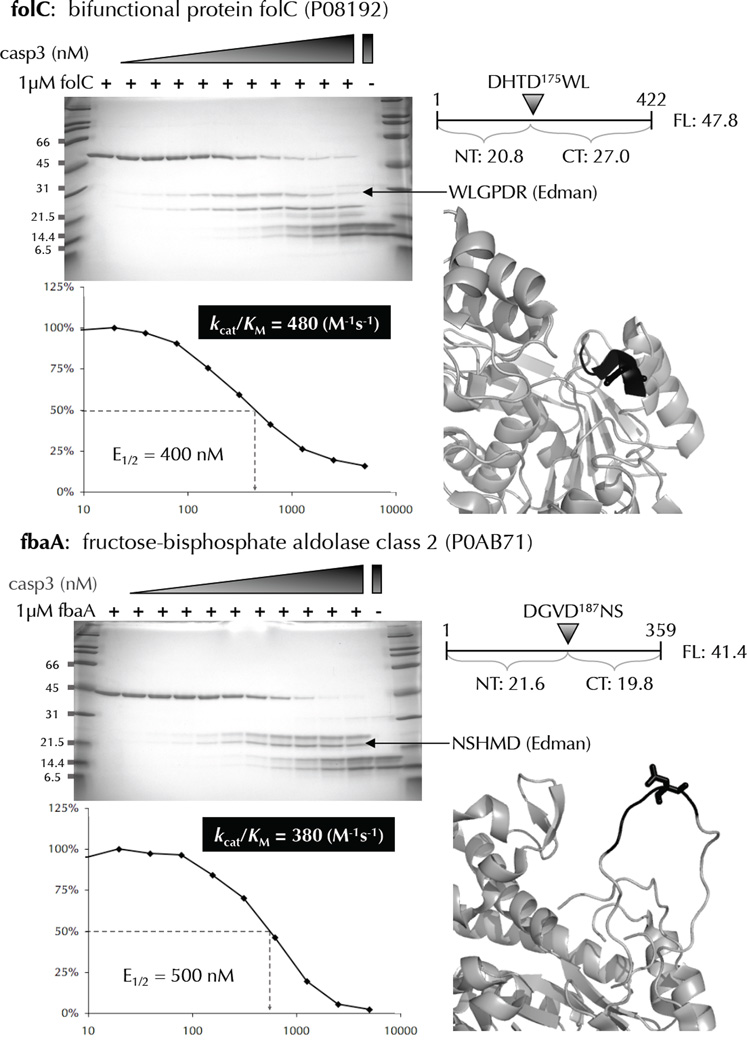
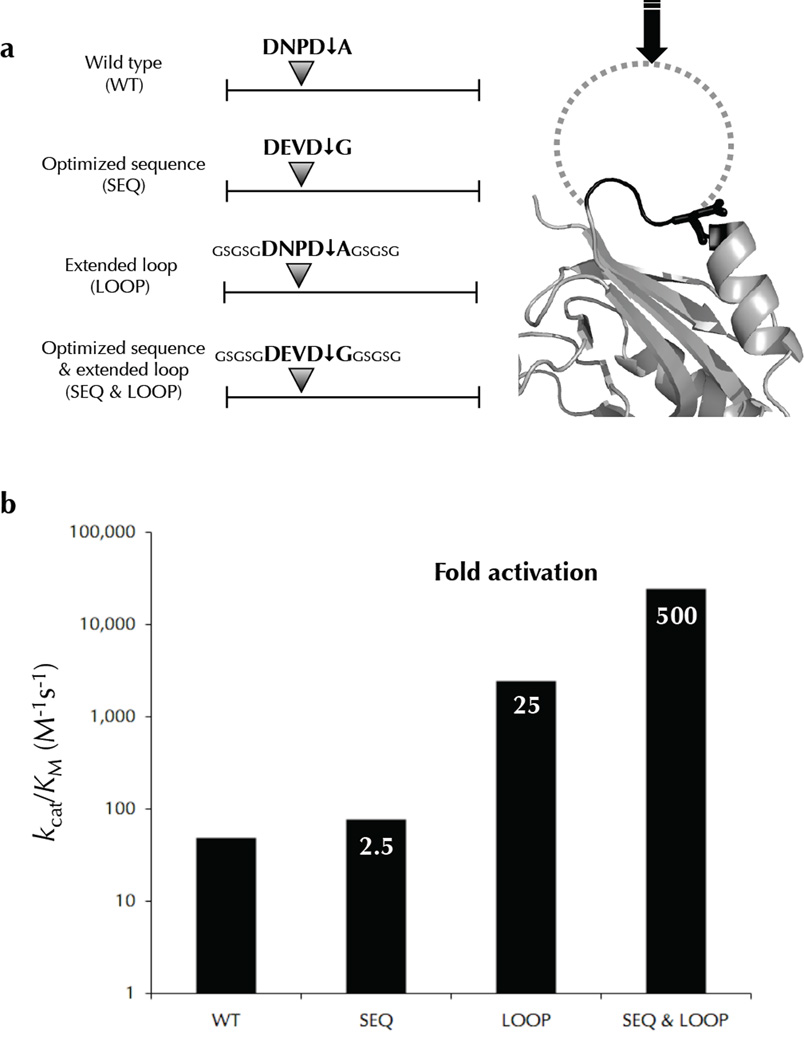
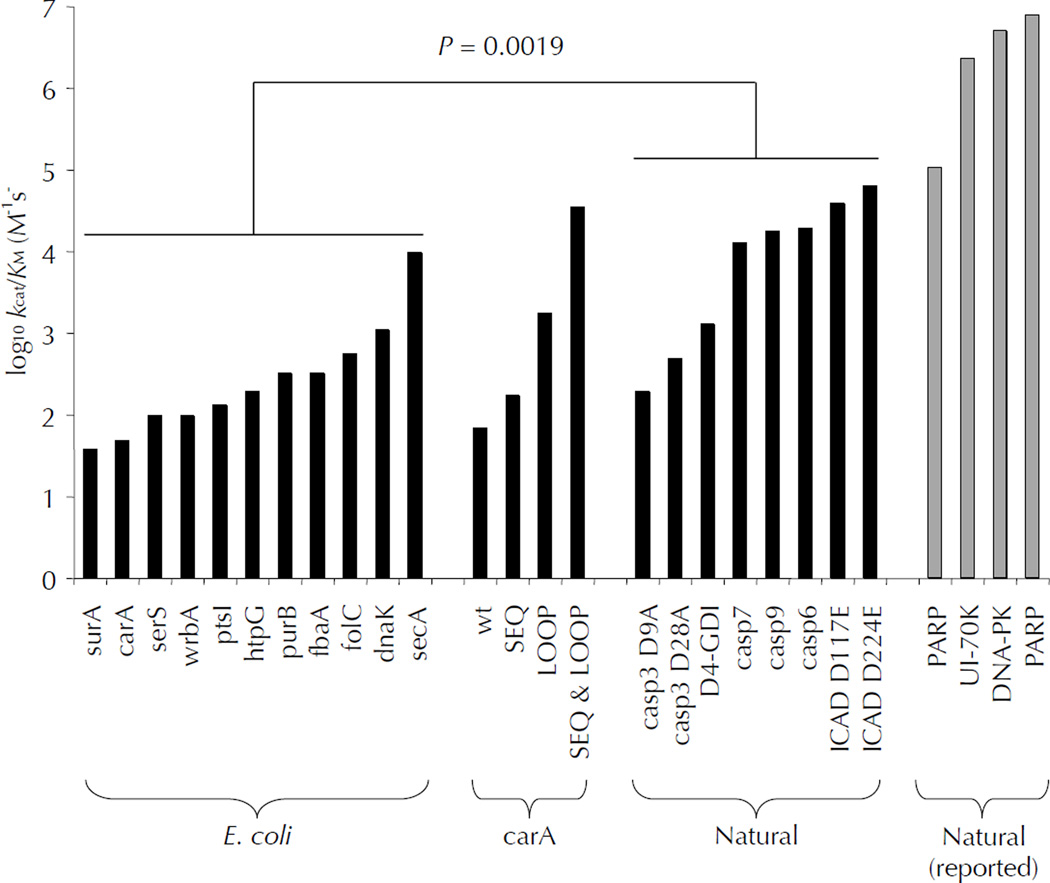
Two fundamental questions with regard to proteolytic networks and pathways concern the structural repertoire and kinetic threshold that distinguish legitimate signaling substrates. We used N-terminal proteomics to address these issues by identifying cleavage sites within the Escherichia coli proteome that are driven by the apoptotic signaling protease caspase-3 and the bacterial protease glutamyl endopeptidase (GluC). Defying the dogma that proteases cleave primarily in natively unstructured loops, we found that both caspase-3 and GluC cleave in alpha-helices nearly as frequently as in extended loops. Notably, biochemical and kinetic characterization revealed that E. coli caspase-3 substrates are greatly inferior to natural substrates, suggesting protease and substrate coevolution. Engineering an E. coli substrate to match natural catalytic rates defined a kinetic threshold that depicts a signaling event. This unique combination of proteomics, biochemistry, kinetics and substrate engineering reveals new insights into the structure-function relationship of protease targets and their validation from large-scale approaches.
Figures




References
-
- Puente XS, Sanchez LM, Overall CM, Lopez-Otin C. Human and mouse proteases: a comparative genomic approach. Nat Rev Genet. 2003;4:544–558. - PubMed
-
- Salvesen GS, Abrams JM. Caspase activation - stepping on the gas or releasing the brakes? Lessons from humans and flies. Oncogene. 2004;23:2774–2784. - PubMed
-
- Gevaert K, et al. Applications of diagonal chromatography for proteome-wide characterization of protein modifications and activity-based analyses. Febs J. 2007;274:6277–6289. - PubMed
-
- Timmer JC, Salvesen GS. Caspase substrates. Cell Death Differ. 2007;14:66–72. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
