

The peptidyl-prolyl isomerase Pin1 determines parathyroid hormone mRNA levels and stability in rat models of secondary hyperparathyroidism
- PMID: 19770516
- PMCID: PMC2752082
- DOI: 10.1172/JCI39522
The peptidyl-prolyl isomerase Pin1 determines parathyroid hormone mRNA levels and stability in rat models of secondary hyperparathyroidism
Abstract
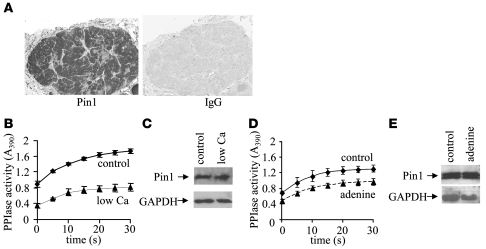
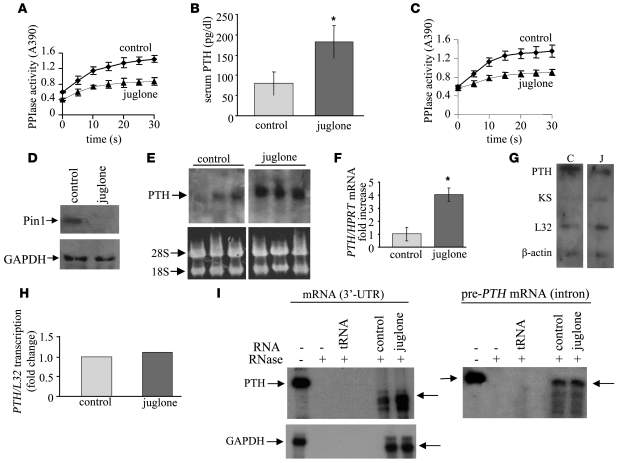
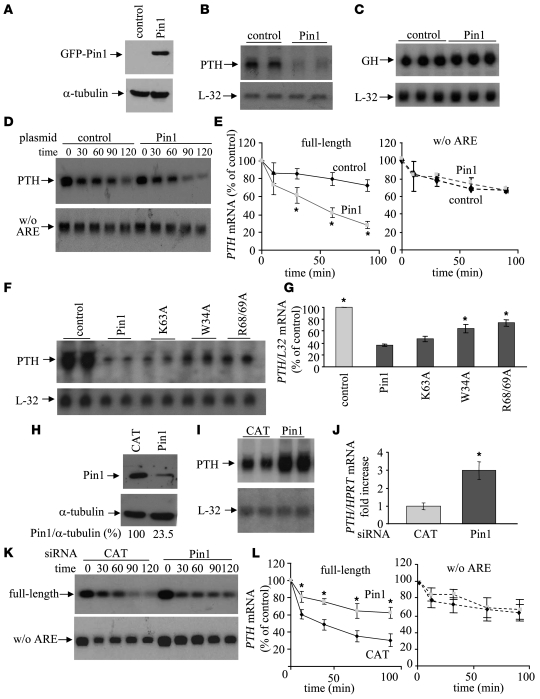
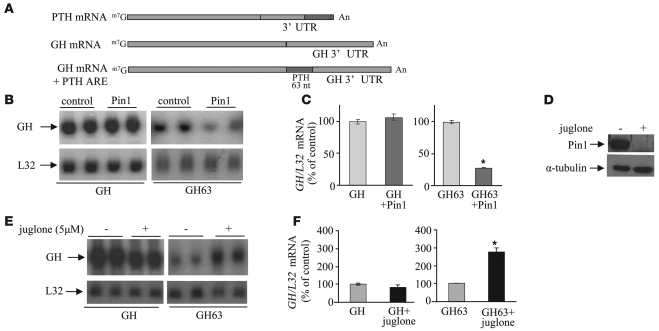
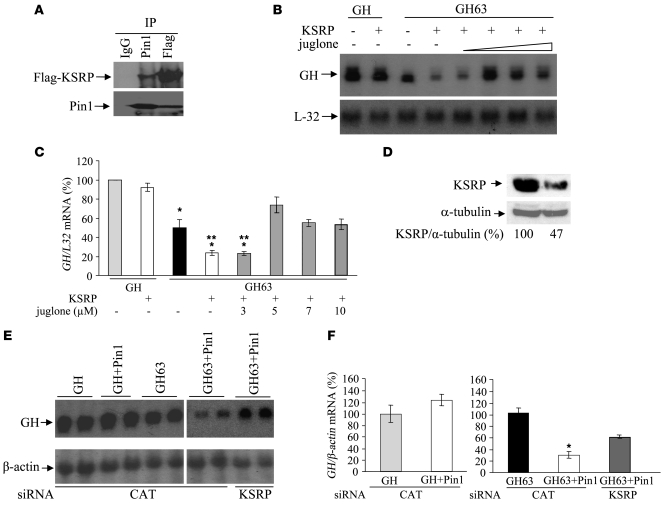
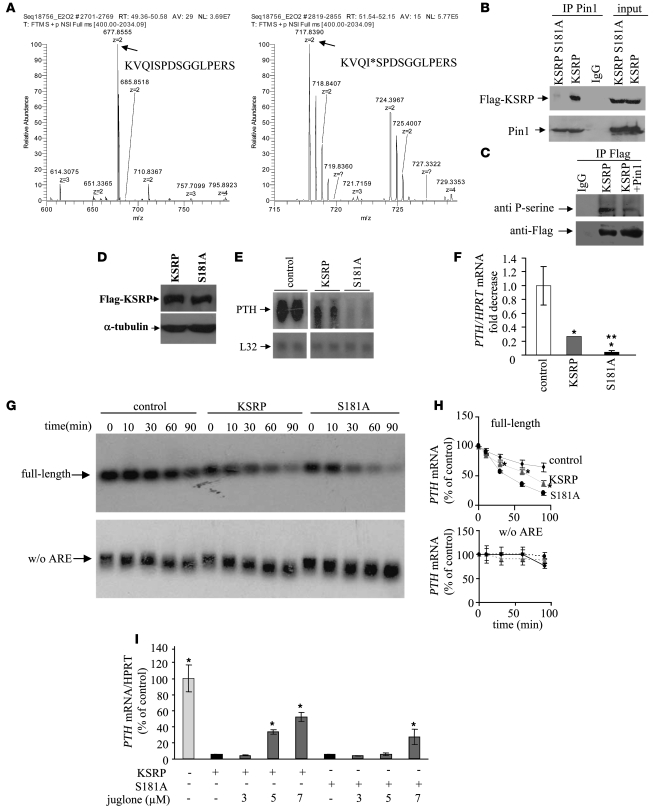
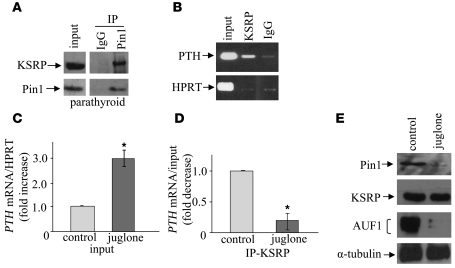
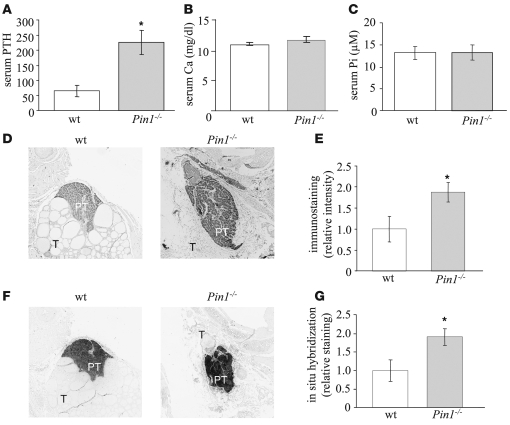
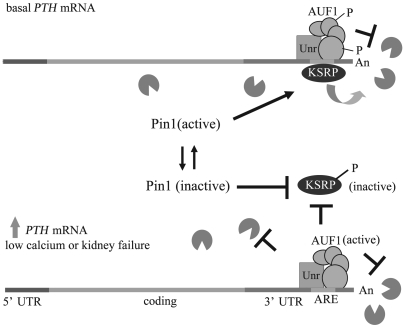
Secondary hyperparathyroidism is a major complication of chronic kidney disease (CKD). In experimental models of secondary hyperparathyroidism induced by hypocalcemia or CKD, parathyroid hormone (PTH) mRNA levels increase due to increased PTH mRNA stability. K-homology splicing regulator protein (KSRP) decreases the stability of PTH mRNA upon binding a cis-acting element in the PTH mRNA 3' UTR region. As the peptidyl-prolyl isomerase (PPIase) Pin1 has recently been shown to regulate the turnover of multiple cytokine mRNAs, we investigated the role of Pin1 in regulating PTH mRNA stability in rat parathyroids and transfected cells. The data generated were consistent with Pin1 being a PTH mRNA destabilizing protein. Initial analysis indicated that Pin1 activity was decreased in parathyroid protein extracts from both hypocalcemic and CKD rats and that pharmacologic inhibition of Pin1 increased PTH mRNA levels posttranscriptionally in rat parathyroid and in transfected cells. Pin1 mediated its effects via interaction with KSRP, which led to KSRP dephosphorylation and activation. In the rat parathyroid, Pin1 inhibition decreased KSRP-PTH mRNA interactions, increasing PTH mRNA levels. Furthermore, Pin1-/- mice displayed increased serum PTH and PTH mRNA levels, suggesting that Pin1 determines basal PTH expression in vivo. These results demonstrate that Pin1 is a key mediator of PTH mRNA stability and indicate a role for Pin1 in the pathogenesis of secondary hyperparathyroidism in individuals with CKD.
Figures
Comment in
-
Pin1 regulates parathyroid hormone mRNA stability.J Clin Invest. 2009 Oct;119(10):2887-91. doi: 10.1172/JCI40784. Epub 2009 Sep 21. J Clin Invest. 2009. PMID: 19770518 Free PMC article.
References
-
- Silver, J., Naveh-Many, T., and Kronenberg, H.M. 2002. Parathyroid hormone: molecular biology. InPrinciples of bone biology . J.B. Bilezikian, L.G. Raisz, and G.A. Rodan, editors. Academic Press. San Diego, California, USA. 407–422.
-
- Levi R., et al. Increased parathyroid hormone gene expression in secondary hyperparathyroidism of experimental uremia is reversed by calcimimetics: correlation with posttranslational modification of the trans acting factor AUF1. J. Am. Soc. Nephrol. 2006;17:107–112. doi: 10.1681/ASN.2005070679. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
