

Fas stimulation of T lymphocytes promotes rapid intercellular exchange of death signals via membrane nanotubes
- PMID: 19770844
- PMCID: PMC2822704
- DOI: 10.1038/cr.2009.112
Fas stimulation of T lymphocytes promotes rapid intercellular exchange of death signals via membrane nanotubes
Abstract
The Fas/CD95 surface receptor mediates rapid death of various cell types, including autoreactive T cells with the potential for triggering autoimmunity. Here, we present novel aspects of Fas signalling that define a 'social' dimension to receptor-induced apoptosis. Fas stimulation rapidly induces extensive membrane nanotube formation between neighbouring T cells. This is critically dependent on Rho GTPases but not on caspase activation. Bidirectional transfer of membrane and cytosolic elements including active caspases can be observed to occur via these nanotubes. Nanotube formation and intercellular exchanges of death signals are defective in T lymphocytes from patients with autoimmune lymphoproliferative syndrome harbouring mutations in the Fas receptor. We conclude that nanotube-mediated exchanges constitute a novel form of intercellular communication that augments the propagation of death signalling between neighbouring T cells.
Figures
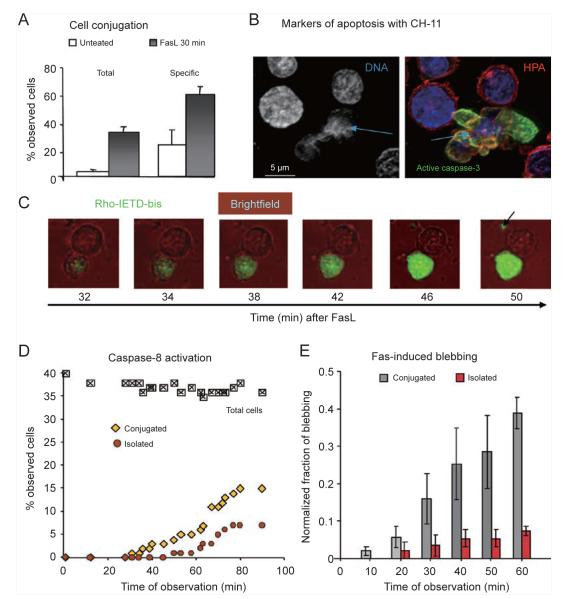
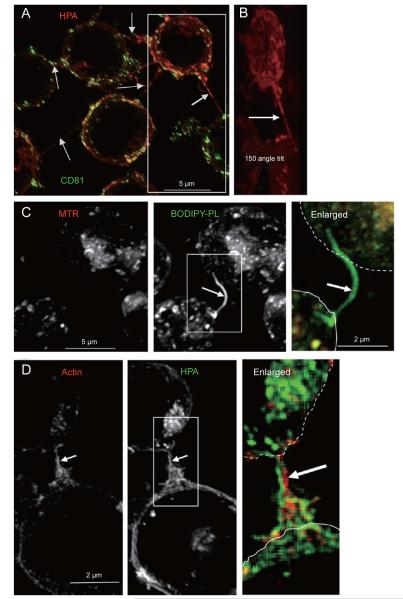
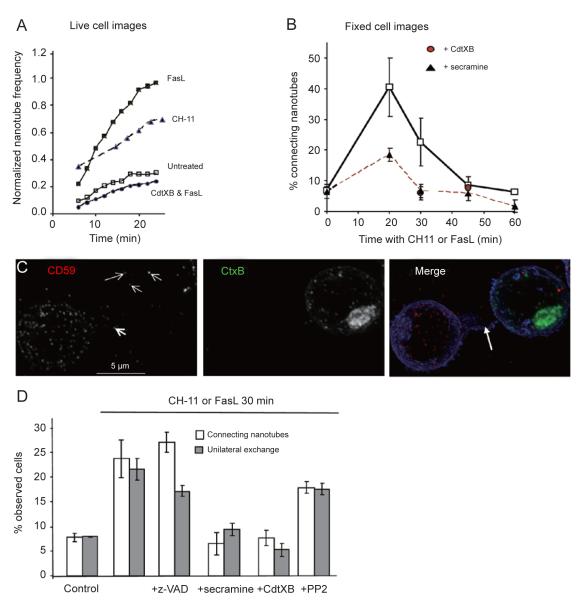
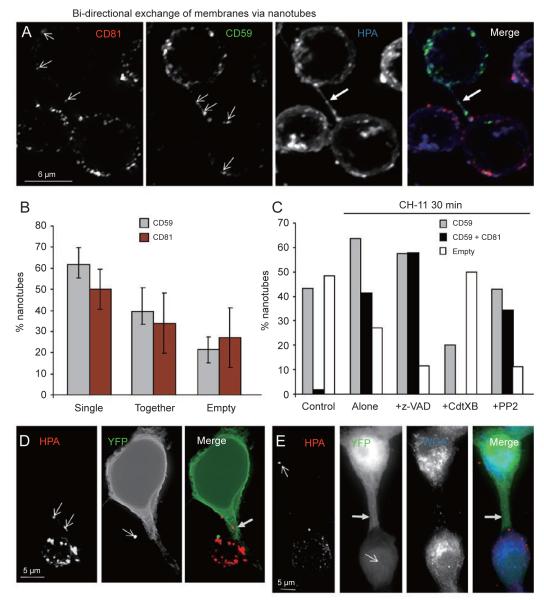
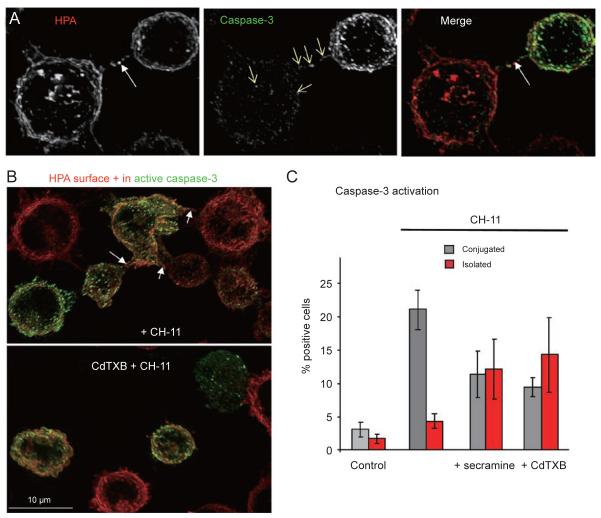
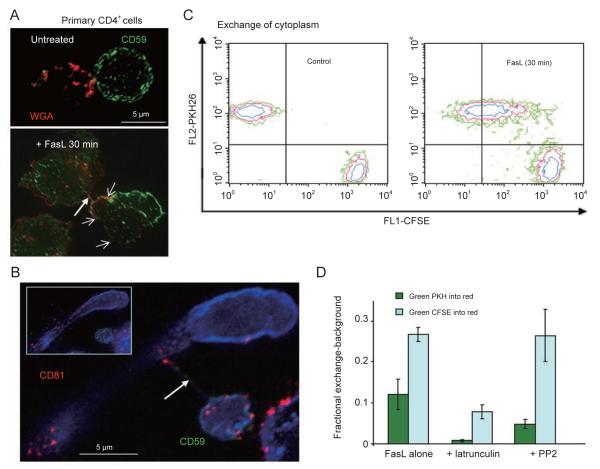
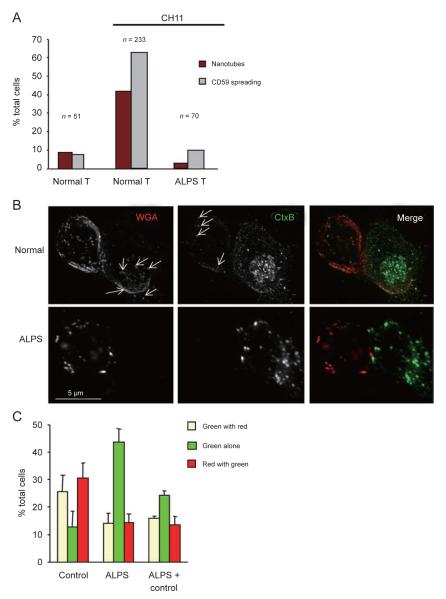
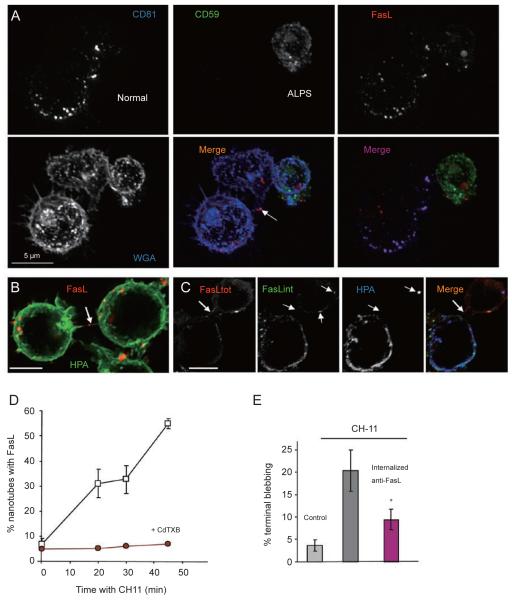
References
-
- Green DR, Droin N, Pinkoski M. Activation-induced cell death in T cells. Immunol Rev. 2003;193:70–81. - PubMed
-
- Peter ME, Krammer PH. The CD95(APO-1/Fas) DISC and beyond. Cell Death Differ. 2003;10:26–35. - PubMed
-
- Rao VK, Straus SE. Causes and consequences of the auto-immune lymphoproliferative syndrome. Hematology. 2006;11:15–23. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
