

Review
doi: 10.1038/nrm2777.
Epub 2009 Sep 23.
Mechanisms of alternative splicing regulation: insights from molecular and genomics approaches
Affiliations
- PMID: 19773805
- PMCID: PMC2958924
- DOI: 10.1038/nrm2777
Item in Clipboard
Review
Mechanisms of alternative splicing regulation: insights from molecular and genomics approaches
Nat Rev Mol Cell Biol.
2009 Nov.
Abstract
Alternative splicing of mRNA precursors provides an important means of genetic control and is a crucial step in the expression of most genes. Alternative splicing markedly affects human development, and its misregulation underlies many human diseases. Although the mechanisms of alternative splicing have been studied extensively, until the past few years we had not begun to realize fully the diversity and complexity of alternative splicing regulation by an intricate protein-RNA network. Great progress has been made by studying individual transcripts and through genome-wide approaches, which together provide a better picture of the mechanistic regulation of alternative pre-mRNA splicing.
Figures
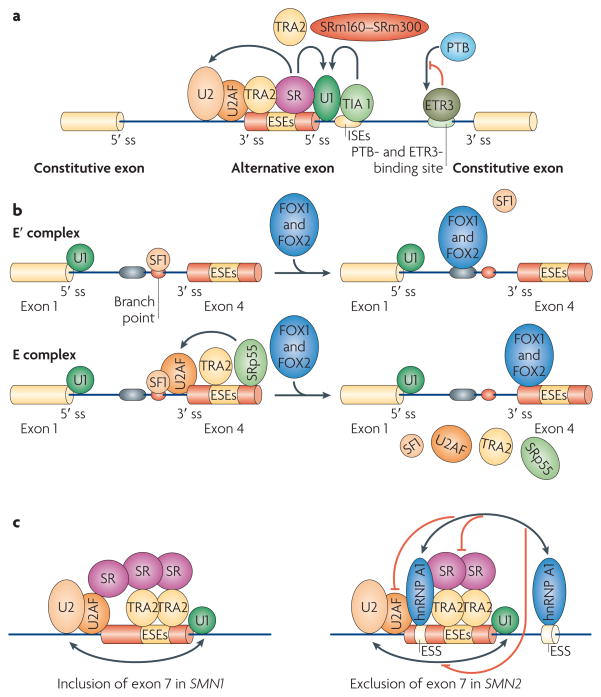
Schematic depicting mechanisms of splicing activation. a | SR (Ser–Arg) proteins bind to exonic splicing enhancers (ESEs) to stimulate the binding of U2AF to the upstream 3′ splice site (ss) or the binding of the U1 small nuclear ribonucleoprotein (snRNP) to the downstream 5′ ss. SR proteins function with other splicing co-activators, such as transformer 2 (TRA2) and the SR-related nuclear matrix proteins SRm160–SRm300. T cell-restricted intracellular antigen 1 (TIA1) binds to U-rich sequences (intronic splicing enhancers (ISEs)) immediately downstream of 5′ splice sites to facilitate U1 binding. CELF (CUGBP- and ETR3-like factor) proteins, such as ETR3, bind to similar sequences as polypyrimidine-tract binding protein (PTB), thereby activating splicing by competing with PTB. b | Fox1 and Fox2 inhibit the inclusion of CALCA (calcitonin-related polypeptide-α) exon 4 by blocking the binding of splicing factor 1 (SF1) to the branch point (top panel) and of TRA2 and SRp55 to ESEs (bottom panel), thereby inhibiting spliceosome assembly at two stages, the E′ and E complexes. The arrow indicates that SRp55 and TRA2 promote binding of the U2AF complex. c | Single-nucleotide differences in SMN2 (survival of motor neuron 2) compared with SMN1 create binding sites for heterogeneous nuclear ribonucleoprotein particle A1 (hnRNP A1) (or hnRNP A2) in exon 7 and in the downstream intron in SMN2 pre-mRNA. hnRNP A1 (or hnRNP A2) may then inhibit the formation or stabilization of the U2 snRNP complex, either directly or indirectly by blocking the activity of the downstream TRA2-dependent ESE. Note that it has also been suggested that the base change in exon 7 destroys an ASF/SF2-dependent ESE, (but see also REF. 69).
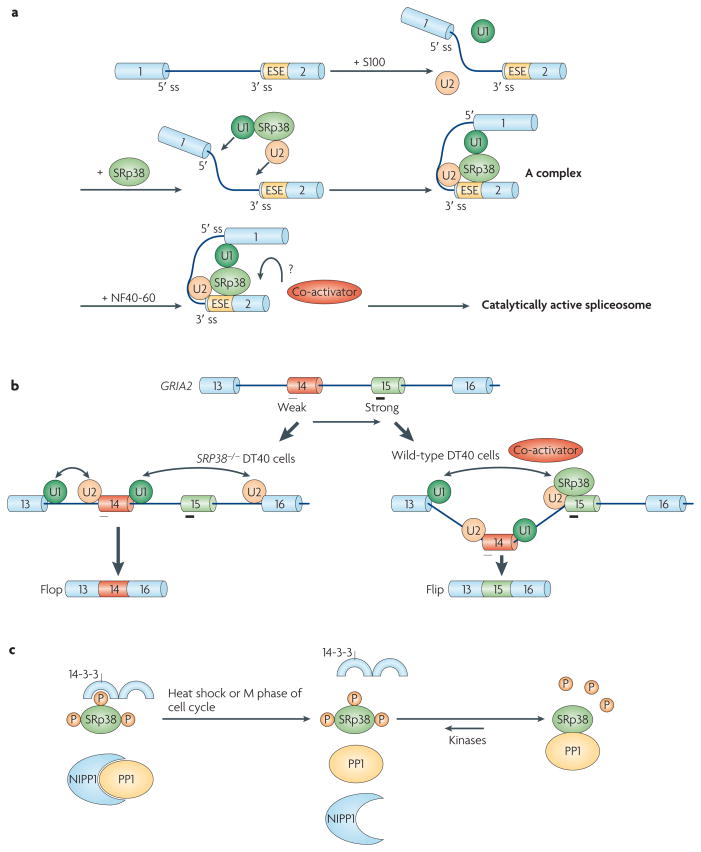
a | Phosphorylated SRp38 (Ser–Arg protein 38) activates splicing by recruiting the U1 and U2 small nuclear ribonucleoprotein particles (snRNPs) to splice sites (ss). SRp38 binds SRp38-dependent exonic splicing enhancers (ESEs) in target transcripts and facilitates the association of U1 and U2 snRNPs with the pre-mRNA to stabilize 5′ ss and branch site recognition by interacting with U1 and U2 snRNPs, respectively. However, the spliceosomal A complex formed is stalled in S100 extract, in which an SRp38-specific cofactor from NF40-60 is absent, which is required to proceed through the splicing pathway. b | SRp38 enhances the inclusion of the Flip exon of GRIA2 (glutamate receptor, ionotropic, AMPA 2) pre-mRNA relative to the mutually exclusive Flop exon. Both exons contain SRp38-binding sites (indicated by black bars under exon 14 (Flop) and exon 15 (Flip)), but the site in Flip is stronger (indicated by the thicker bar), and Flip inclusion is therefore favoured in the presence of SRp38. c | Protein phosphatase 1 (PP1) dephosphorylates SRp38 on heat shock. Under normal conditions, phosphorylated SRp38 is associated with 14-3-3 proteins, which help to protect SRp38 from dephosphorylation, and PP1 activity is inhibited by PP1-associated proteins, including nuclear inhibitor of PP1 (NIPP1). During heat shock, PP1 dissociates from NIPP1 and directly binds to and dephosphorylates SRp38, which has dissociated from 14-3-3 proteins. Part a of the figure modified, with permission, from Nature Struct. Mol. Biol. REF. © (2008) Macmillan Publishers Ltd. All rights reserved. Part c of the figure modified, with permission, from REF. © (2007) Elsevier.
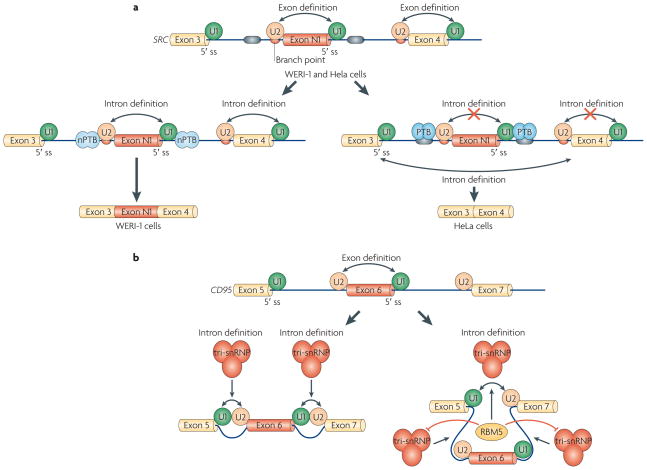
a | Polypyrimidine-tract binding protein (PTB) inhibits the inclusion of SRC exon N1 by inhibiting the interactions of the U1 and U2 small nuclear ribonucleoprotein particles (snRNPs) and intron definition. In both WERI-1 and HeLa cells, the N1 exon is defined by the binding of U1 snRNP to the 5′ splice site (ss) and of U2 snRNP to the branch point. In WERI-1 cells, in the absence of PTB, U1 and U2 snRNPs bound to the N1 exon interact with the U2 and U1 snRNP on adjacent constitutive exons, respectively, thereby allowing efficient spliceosome assembly on introns flanking exon N1. In HeLa cells, PTB binds to sequences flanking exon N1 and prevents the cross-intron interactions that occur in WERI-cells, thereby excluding exon N1. b | RBM5 (RNA-binding protein 5) regulates alternative splicing of CD95 by inhibiting the inclusion of exon 6. It does so not at the stage of U1 and U2 snRNP binding, but instead by promoting tri-snRNP assembly on the intron-defined spliceosomal complex between exon 5 and exon 7, while blocking tri-snRNP recruitment to complexes that would result in inclusion of exon 6. Double-headed arrows indicate intron and exon definition.
References
-
- Black DL. Mechanisms of alternative pre-messenger RNA splicing. Annu Rev Biochem. 2003;72:291–336. - PubMed
-
- Wahl MC, Will CL, Luhrmann R. The spliceosome: design principles of a dynamic RNP machine. Cell. 2009;136:701–718. - PubMed
-
- Berglund JA, Chua K, Abovich N, Reed R, Rosbash M. The splicing factor BBP interacts specifically with the pre-mRNA branchpoint sequence UACUAAC. Cell. 1997;89:781–787. - PubMed
-
- Nelson KK, Green MR. Mammalian U2 snRNP has a sequence-specific RNA-binding activity. Genes Dev. 1989;3:1562–1571. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
