

Review
doi: 10.1016/j.ceb.2009.08.004.
Epub 2009 Sep 21.
Control of cell growth by the SCF and APC/C ubiquitin ligases
Affiliations
- PMID: 19775879
- PMCID: PMC2805079
- DOI: 10.1016/j.ceb.2009.08.004
Item in Clipboard
Review
Control of cell growth by the SCF and APC/C ubiquitin ligases
Curr Opin Cell Biol.
2009 Dec.
Abstract
The ubiquitin-proteasome system plays key roles in the control of cell growth. The cell cycle, in particular, is highly regulated by the functions of the SCF and APC/C ubiquitin ligases, and perturbation of their function can result in tumorigenesis. Although the SCF and APC/C complexes are well established in growth control pathways, many aspects of their function remain unknown. Recent studies have shed light on the mechanism of SCF-mediated ubiquitination and new functions for the SCF complex and APC/C. Our expanding understanding of the roles of the SCF and APC/C complexes highlight the potential for targeted molecular therapies.
Figures

Non-neddylated CUL1 holds RBX1 in a closed form and is bound by CAND1, preventing SKP1-F-box protein binding. Neddylation of CUL1 releases RBX1 on a flexible linker and dissociates CAND1, allowing binding of the SKP1-F-box protein pair. The flexible RBX1 linker bridges the gap between the E2 and substrate (that would be present in non-neddylated SCF), facilitating both the initial ubiquitination and subsequent polyubiquitination.
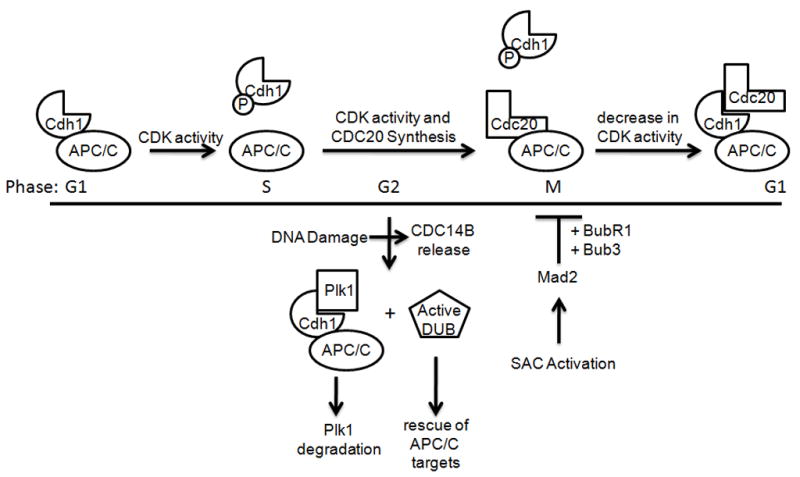
APC/C is activity is highly regulated during the cell cycle through the binding of the Cdc20 and Cdh1 activating subunits. APC/CCdh1 is active in G1, but Cdh1 is inactivated by increasing amounts of Cdk activity as the cell cycle progresses, preventing binding to the APC/C core. (Other means of Cdh1 inactivation, including Emi1 binding and degradation are not depicted here.) Cdc20 levels are low in G1 due to APC/CCdh1-mediated degradation, but they increase through S and G2. In M, APC/CCdc20 forms, but it remains inactive until bipolar attachment of all chromosomes to the spindle and the Spindle Assembly Checkpoint (SAC) is satisfied. Then, APC/CCdc20 targets Securin, Shugoshin, Cyclin B, and other proteins for degradation. With decreasing Cdk activity and Cdc14 phosphatase activity, unphosphorylated Cdh1 binds the APC/C core, resulting in the targeting of Cdc20 and other substrates (many of which are also APC/CCdc20 substrates) whose degradation is required to exit mitosis, enter G1, and maintain the G1 state. Although Cdh1 is inactive due to phosphorylation in G2, DNA damage releases the Cdc14B phosphatase from the nucleolus, causing Cdh1 dephosphorylation, resulting in active APC/CCdh1, which targets Plk1 for degradation. Notably, certain APC/CCdh1 substrates, such as Claspin, are spared, likely through the action of deubiquitinating enzymes (DUBs).

β-TRCP controls early events in the cell cycle, such as the response to mitogens, but it also controls later events in the cell cycle through Cdk1, the APC/C, and Plk1. Notably, feedback loops are common in the control of the Cdk1 and Plk1 kinases. During DNA damage responses, β-TRCP targets for Cdc25A for degradation, but during the recovery from DNA damage, β-TRCP targets Clapsin for degradation, blocking Chk1 activation and Cdc25A degradation. Additionally, Plk1 is involved in the degradation of multiple β-TRCP substrates, and Bora, which activates Plk1, is targeted for β-TRCP-dependent degradation by Plk1, attenuating Plk1 activity at the end of mitosis. Notably, there is also much crosstalk between β-TRCP targets, with APC/C activity controlled by Cdk1 (via phosphorylation) and Plk1 (via phosphorylation and induction of Emi1 degradation). At the same time, APC/C is required for the inactivation of Cdk1 and Plk1 activators (cyclins and Bora, respectively). Finally, Plk1 is a positive regulator of Cdk1 through the induction of Claspin and Wee1 degradation.
References
-
- Pickart CM, Fushman D. Polyubiquitin chains: polymeric protein signals. Curr Opin Chem Biol. 2004;8:610–616. - PubMed
-
- Hershko A, Ciechanover A. The ubiquitin system. Annu Rev Biochem. 1998;67:425–479. - PubMed
-
- Schwartz AL, Ciechanover A. The ubiquitin-proteasome pathway and pathogenesis of human diseases. Annu Rev Med. 1999;50:57–74. - PubMed
-
- Weissman AM. Themes and variations on ubiquitylation. Nat Rev Mol Cell Biol. 2001;2:169–178. - PubMed
-
- Cardozo T, Pagano M. The SCF ubiquitin ligase: insights into a molecular machine. Nat Rev Mol Cell Biol. 2004;5:739–751. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
