

Crystal structure and functional analysis of homocitrate synthase, an essential enzyme in lysine biosynthesis
- PMID: 19776021
- PMCID: PMC2791007
- DOI: 10.1074/jbc.M109.046821
Crystal structure and functional analysis of homocitrate synthase, an essential enzyme in lysine biosynthesis
Abstract
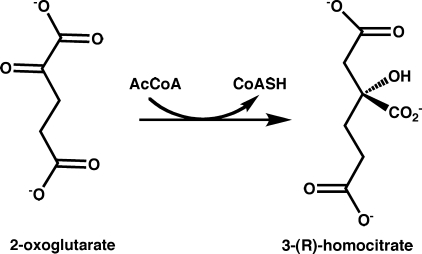
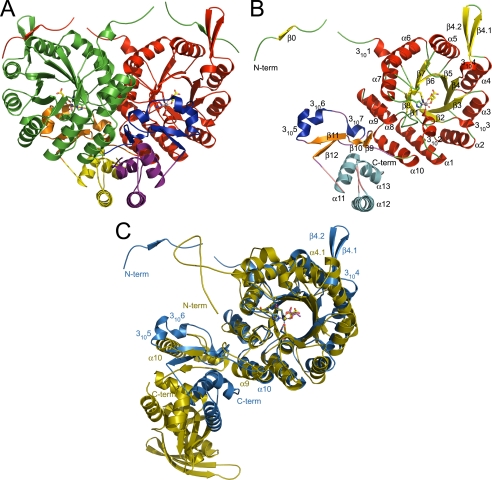
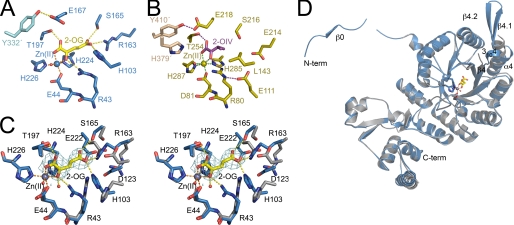
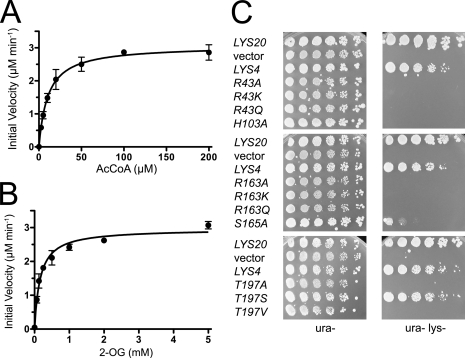
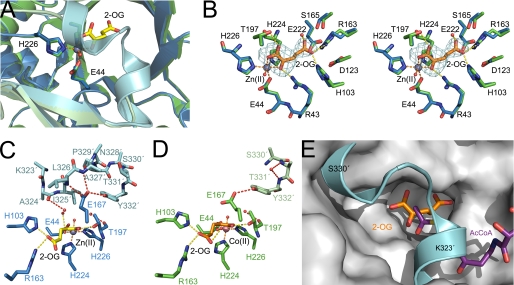
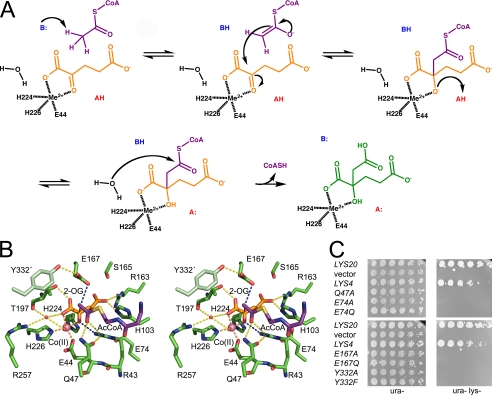
Homocitrate synthase (HCS) catalyzes the first and committed step in lysine biosynthesis in many fungi and certain Archaea and is a potential target for antifungal drugs. Here we report the crystal structure of the HCS apoenzyme from Schizosaccharomyces pombe and two distinct structures of the enzyme in complex with the substrate 2-oxoglutarate (2-OG). The structures reveal that HCS forms an intertwined homodimer stabilized by domain-swapping between the N- and C-terminal domains of each monomer. The N-terminal catalytic domain is composed of a TIM barrel fold in which 2-OG binds via hydrogen bonds and coordination to the active site divalent metal ion, whereas the C-terminal domain is composed of mixed alpha/beta topology. In the structures of the HCS apoenzyme and one of the 2-OG binary complexes, a lid motif from the C-terminal domain occludes the entrance to the active site of the neighboring monomer, whereas in the second 2-OG complex the lid is disordered, suggesting that it regulates substrate access to the active site through its apparent flexibility. Mutations of the active site residues involved in 2-OG binding or implicated in acid-base catalysis impair or abolish activity in vitro and in vivo. Together, these results yield new insights into the structure and catalytic mechanism of HCSs and furnish a platform for developing HCS-selective inhibitors.
Figures
Similar articles
-
Structural basis for L-lysine feedback inhibition of homocitrate synthase.J Biol Chem. 2010 Apr 2;285(14):10446-53. doi: 10.1074/jbc.M109.094383. Epub 2010 Jan 19. J Biol Chem. 2010. PMID: 20089861 Free PMC article.
-
Mechanism of substrate recognition and insight into feedback inhibition of homocitrate synthase from Thermus thermophilus.J Biol Chem. 2010 Feb 5;285(6):4195-4205. doi: 10.1074/jbc.M109.086330. Epub 2009 Dec 7. J Biol Chem. 2010. PMID: 19996101 Free PMC article.
-
Involvement of subdomain II in the recognition of acetyl-CoA revealed by the crystal structure of homocitrate synthase from Sulfolobus acidocaldarius.FEBS J. 2021 Mar;288(6):1975-1988. doi: 10.1111/febs.15527. Epub 2020 Sep 8. FEBS J. 2021. PMID: 32897601
-
Inactivation of homocitrate synthase causes lysine auxotrophy in copper/zinc-containing superoxide dismutase-deficient yeast Schizosaccharomyces pombe.J Biol Chem. 2006 Jan 20;281(3):1345-51. doi: 10.1074/jbc.M506611200. Epub 2005 Nov 18. J Biol Chem. 2006. PMID: 16299000
-
High-Level Production of Lysine in the Yeast Saccharomyces cerevisiae by Rational Design of Homocitrate Synthase.Appl Environ Microbiol. 2021 Jul 13;87(15):e0060021. doi: 10.1128/AEM.00600-21. Epub 2021 Jul 13. Appl Environ Microbiol. 2021. PMID: 33990312 Free PMC article.
Cited by
-
Identification and characterization of re-citrate synthase in Syntrophus aciditrophicus.J Bacteriol. 2013 Apr;195(8):1689-96. doi: 10.1128/JB.02185-12. Epub 2013 Feb 1. J Bacteriol. 2013. PMID: 23378508 Free PMC article.
-
Structural basis for L-lysine feedback inhibition of homocitrate synthase.J Biol Chem. 2010 Apr 2;285(14):10446-53. doi: 10.1074/jbc.M109.094383. Epub 2010 Jan 19. J Biol Chem. 2010. PMID: 20089861 Free PMC article.
-
Application of a high-throughput fluorescent acetyltransferase assay to identify inhibitors of homocitrate synthase.Anal Biochem. 2011 Mar 1;410(1):133-40. doi: 10.1016/j.ab.2010.11.004. Epub 2010 Nov 10. Anal Biochem. 2011. PMID: 21073853 Free PMC article.
-
New Therapeutic Candidates for the Treatment of Malassezia pachydermatis -Associated Infections.Sci Rep. 2020 Mar 17;10(1):4860. doi: 10.1038/s41598-020-61729-1. Sci Rep. 2020. PMID: 32184419 Free PMC article.
-
A moonlighting metabolic protein influences repair at DNA double-stranded breaks.Nucleic Acids Res. 2015 Feb 18;43(3):1646-58. doi: 10.1093/nar/gku1405. Epub 2015 Jan 27. Nucleic Acids Res. 2015. PMID: 25628362 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
