

The mammalian ovary from genesis to revelation
- PMID: 19776209
- PMCID: PMC2761115
- DOI: 10.1210/er.2009-0012
The mammalian ovary from genesis to revelation
Abstract
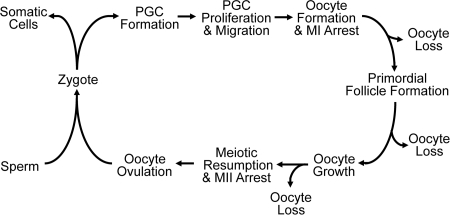
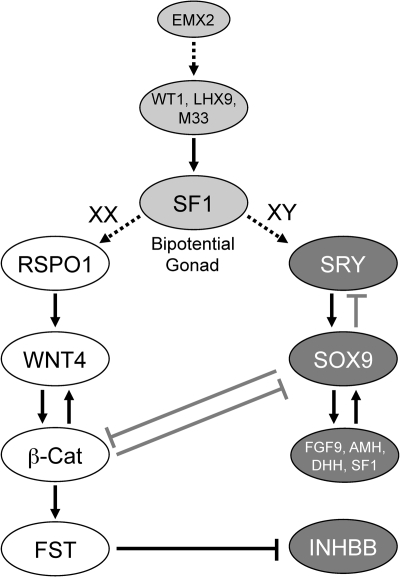
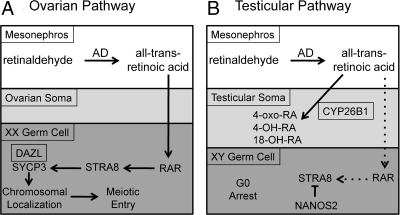
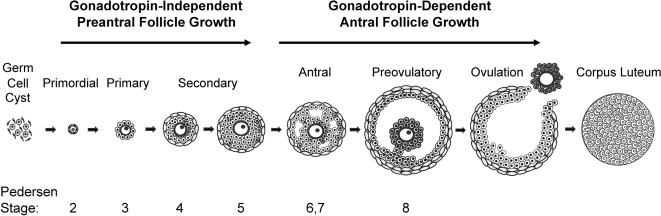
Two major functions of the mammalian ovary are the production of germ cells (oocytes), which allow continuation of the species, and the generation of bioactive molecules, primarily steroids (mainly estrogens and progestins) and peptide growth factors, which are critical for ovarian function, regulation of the hypothalamic-pituitary-ovarian axis, and development of secondary sex characteristics. The female germline is created during embryogenesis when the precursors of primordial germ cells differentiate from somatic lineages of the embryo and take a unique route to reach the urogenital ridge. This undifferentiated gonad will differentiate along a female pathway, and the newly formed oocytes will proliferate and subsequently enter meiosis. At this point, the oocyte has two alternative fates: die, a common destiny of millions of oocytes, or be fertilized, a fate of at most approximately 100 oocytes, depending on the species. At every step from germline development and ovary formation to oogenesis and ovarian development and differentiation, there are coordinated interactions of hundreds of proteins and small RNAs. These studies have helped reproductive biologists to understand not only the normal functioning of the ovary but also the pathophysiology and genetics of diseases such as infertility and ovarian cancer. Over the last two decades, parallel progress has been made in the assisted reproductive technology clinic including better hormonal preparations, prenatal genetic testing, and optimal oocyte and embryo analysis and cryopreservation. Clearly, we have learned much about the mammalian ovary and manipulating its most important cargo, the oocyte, since the birth of Louise Brown over 30 yr ago.
Figures
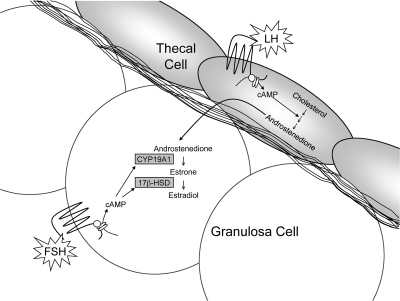
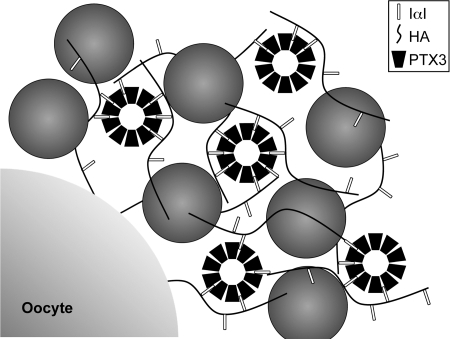
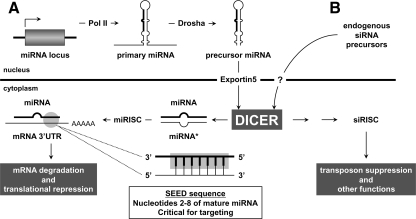
References
-
- Chiquoine AD 1954 The identification, origin and migration of the primordial germ cells in the mouse embryo. Anat Rec 118:135–146 - PubMed
-
- Mintz B, Russell ES 1957 Gene-induced embryological modifications of primordial germ cells in the mouse. J Exp Zool 134:207–237 - PubMed
-
- McCoshen JA, McCallion DJ 1975 A study of the primordial germ cells during their migratory phase in Steel mutant mice. Experientia 31:589–590 - PubMed
-
- Ozdzenski W 1967 Observations on the origin of the primordial germ cells in the mouse. Zool Polon 17:367–379
-
- Spiegelman M, Bennett D 1973 A light- and electron-microscopic study of primordial germ cells in the early mouse embryo. J Embryol Exp Morphol 30:97–118 - PubMed
Publication types
MeSH terms
Grants and funding
- R01 HD033437/HD/NICHD NIH HHS/United States
- U01HD60496/HD/NICHD NIH HHS/United States
- T32 HD007165/HD/NICHD NIH HHS/United States
- U01 HD060496/HD/NICHD NIH HHS/United States
- U54 HD007495/HD/NICHD NIH HHS/United States
- P01 HD036289/HD/NICHD NIH HHS/United States
- T32 GM008307/GM/NIGMS NIH HHS/United States
- P01HD36289/HD/NICHD NIH HHS/United States
- R01CA60651/CA/NCI NIH HHS/United States
- R01 HD032067/HD/NICHD NIH HHS/United States
- R01HD32067/HD/NICHD NIH HHS/United States
- R37HD33437/HD/NICHD NIH HHS/United States
- R01 CA060651/CA/NCI NIH HHS/United States
- K12 DK083014/DK/NIDDK NIH HHS/United States
