

Transformation of polarized light information in the central complex of the locust
- PMID: 19776265
- PMCID: PMC6666666
- DOI: 10.1523/JNEUROSCI.1870-09.2009
Transformation of polarized light information in the central complex of the locust
Abstract
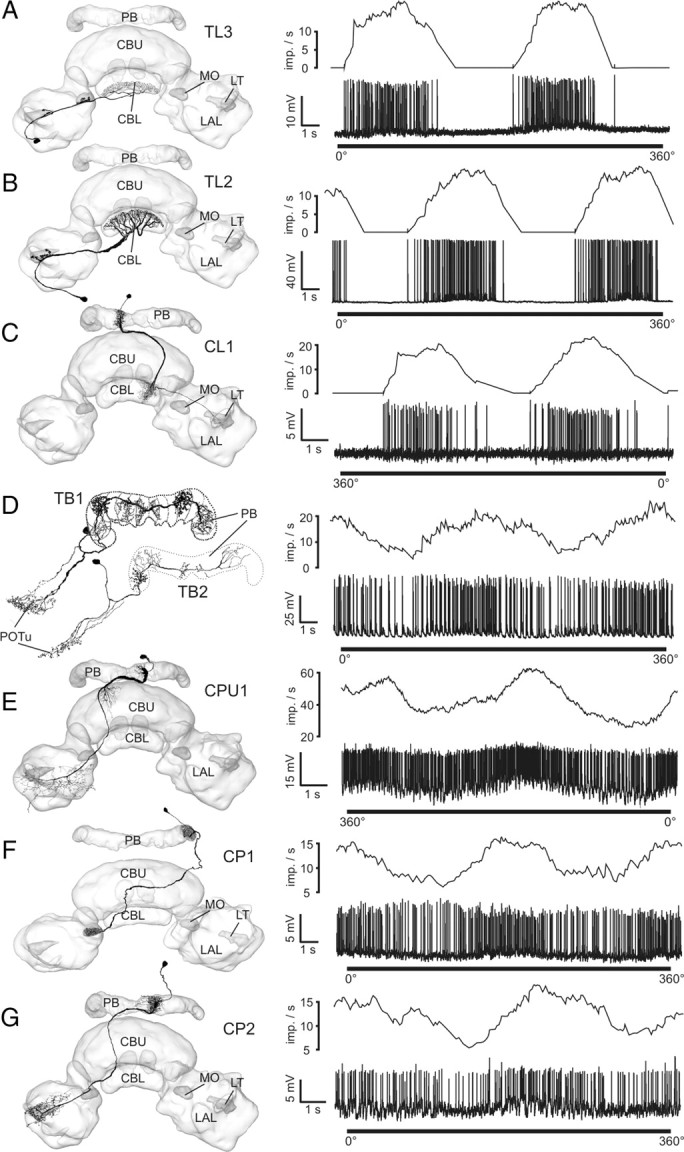
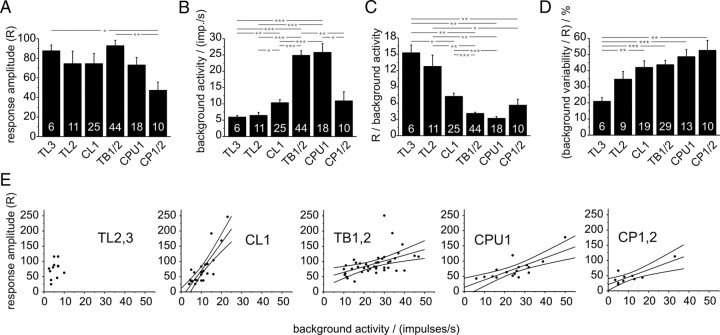
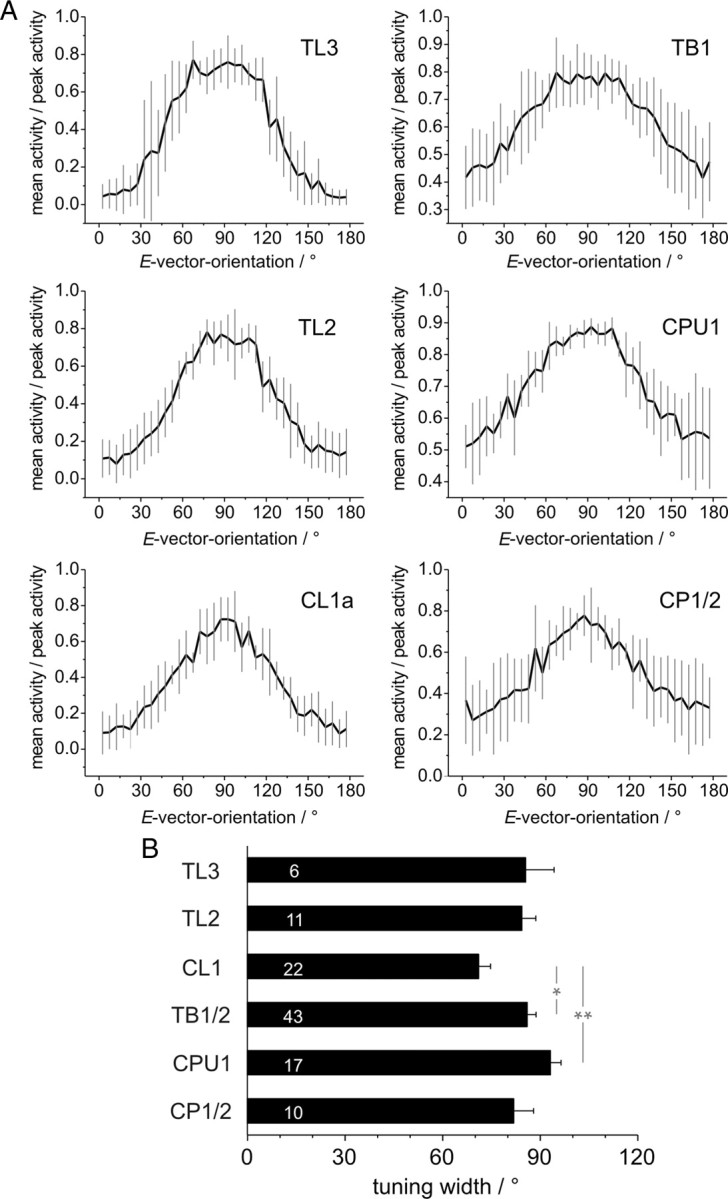
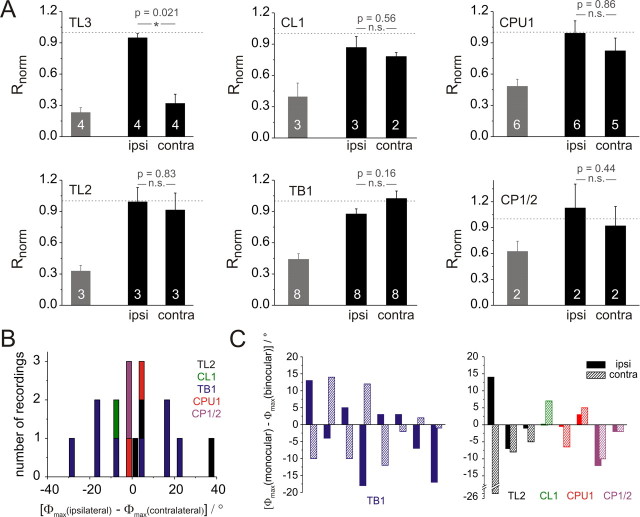
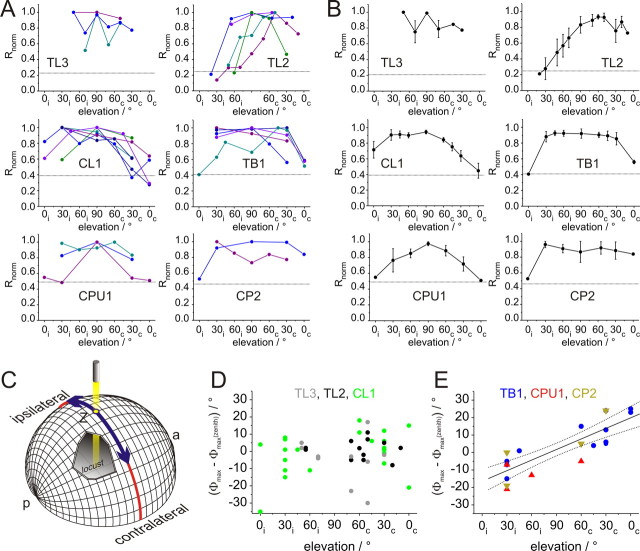
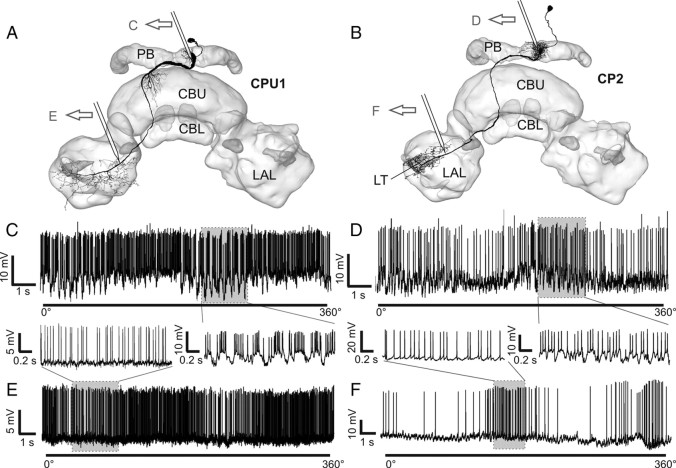
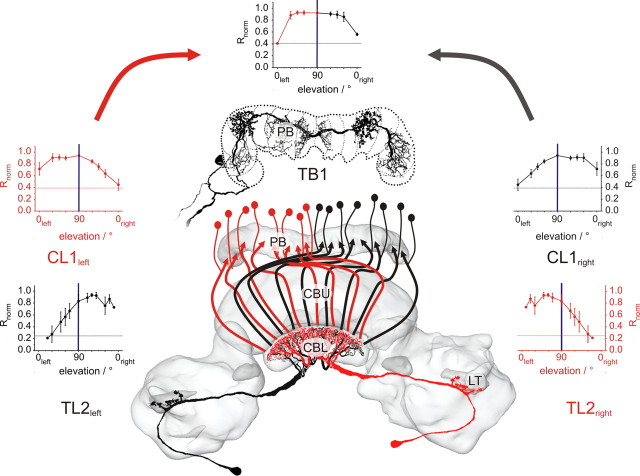
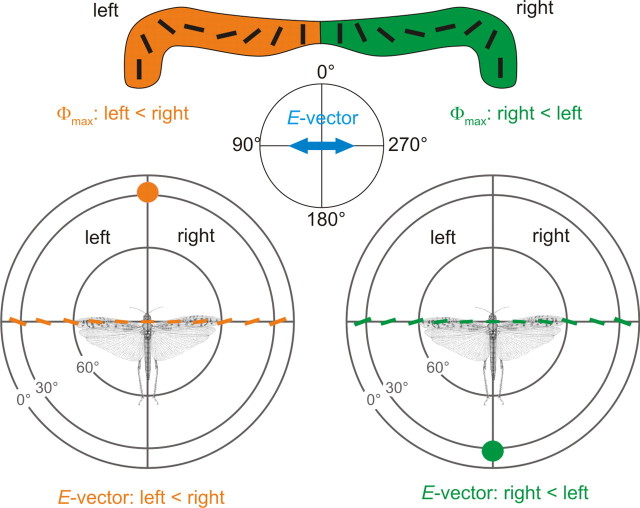
Many insects perceive the E-vector orientation of polarized skylight and use it for compass navigation. In locusts, polarized light is detected by photoreceptors of the dorsal rim area of the eye. Polarized light signals from both eyes are integrated in the central complex (CC), a group of neuropils in the center of the brain. Thirteen types of CC neuron are sensitive to dorsally presented, polarized light (POL-neurons). These neurons interconnect the subdivisions of the CC, particularly the protocerebral bridge (PB), the upper and lower divisions of the central body (CBU, CBL), and the adjacent lateral accessory lobes (LALs). All POL-neurons show polarization-opponency, i.e., receive excitatory and inhibitory input at orthogonal E-vector orientations. To provide physiological evidence for the direction of information flow through the polarization vision network in the CC, we analyzed the functional properties of the different cell types through intracellular recordings. Tangential neurons of the CBL showed highest signal-to-noise ratio, received either ipsilateral polarized-light input only or, together with CL1 columnar neurons, had eccentric receptive fields. Bilateral polarized-light inputs with zenith-centered receptive fields were found in tangential neurons of the PB and in columnar neurons projecting to the LALs. Together with other physiological parameters, these data suggest a flow of information from the CBL (input) to the PB and from here to the LALs (output). This scheme is supported by anatomical data and suggests transformation of purely sensory E-vector coding at the CC input stage to position-invariant coding of 360 degrees -compass directions at the output stage.
Figures
Similar articles
-
Linking the input to the output: new sets of neurons complement the polarization vision network in the locust central complex.J Neurosci. 2009 Apr 15;29(15):4911-21. doi: 10.1523/JNEUROSCI.0332-09.2009. J Neurosci. 2009. PMID: 19369560 Free PMC article.
-
Neurons of the central complex of the locust Schistocerca gregaria are sensitive to polarized light.J Neurosci. 2002 Feb 1;22(3):1114-25. doi: 10.1523/JNEUROSCI.22-03-01114.2002. J Neurosci. 2002. PMID: 11826140 Free PMC article.
-
Neurons in the brain of the desert locust Schistocerca gregaria sensitive to polarized light at low stimulus elevations.J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2016 Nov;202(11):759-781. doi: 10.1007/s00359-016-1116-x. Epub 2016 Aug 3. J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2016. PMID: 27487785
-
In search of the sky compass in the insect brain.Naturwissenschaften. 2004 May;91(5):199-208. doi: 10.1007/s00114-004-0525-9. Epub 2004 Apr 20. Naturwissenschaften. 2004. PMID: 15146265 Review.
-
Central neural coding of sky polarization in insects.Philos Trans R Soc Lond B Biol Sci. 2011 Mar 12;366(1565):680-7. doi: 10.1098/rstb.2010.0199. Philos Trans R Soc Lond B Biol Sci. 2011. PMID: 21282171 Free PMC article. Review.
Cited by
-
Use of bilateral information to determine the walking direction during orientation to a pheromone source in the silkmoth Bombyx mori.J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2012 Apr;198(4):295-307. doi: 10.1007/s00359-011-0708-8. Epub 2012 Jan 8. J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2012. PMID: 22227850
-
Representation of the brain's superior protocerebrum of the flesh fly, Neobellieria bullata, in the central body.J Comp Neurol. 2012 Oct 1;520(14):3070-87. doi: 10.1002/cne.23094. J Comp Neurol. 2012. PMID: 22434505 Free PMC article.
-
Amplitude and dynamics of polarization-plane signaling in the central complex of the locust brain.J Neurophysiol. 2015 May 1;113(9):3291-311. doi: 10.1152/jn.00742.2014. Epub 2015 Jan 21. J Neurophysiol. 2015. PMID: 25609107 Free PMC article.
-
Evolutionarily conserved mechanisms for the selection and maintenance of behavioural activity.Philos Trans R Soc Lond B Biol Sci. 2015 Dec 19;370(1684):20150053. doi: 10.1098/rstb.2015.0053. Philos Trans R Soc Lond B Biol Sci. 2015. PMID: 26554043 Free PMC article.
-
Widespread sensitivity to looming stimuli and small moving objects in the central complex of an insect brain.J Neurosci. 2013 May 8;33(19):8122-33. doi: 10.1523/JNEUROSCI.5390-12.2013. J Neurosci. 2013. PMID: 23658153 Free PMC article.
References
-
- Clements AN, May TE. Studies on locust neuromuscular physiology in relation to glutamic acid. J Exp Biol. 1974;60:673–705. - PubMed
-
- Heinze S, Homberg U. Maplike representation of celestial E-vector orientations in the brain of an insect. Science. 2007;315:995–997. - PubMed
-
- Heinze S, Homberg U. Neuroarchitecture of the central complex of the desert locust: Intrinsic and columnar neurons. J Comp Neurol. 2008;511:454–478. - PubMed
-
- Homberg U. Neuroarchitecture of the central complex in the brain of the locust Schistocerca gregaria and S. americana as revealed by serotonin immunocytochemistry. J Comp Neurol. 1991;303:245–254. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Miscellaneous