

Characterization of small RNAs in Aplysia reveals a role for miR-124 in constraining synaptic plasticity through CREB
- PMID: 19778509
- PMCID: PMC2875683
- DOI: 10.1016/j.neuron.2009.05.029
Characterization of small RNAs in Aplysia reveals a role for miR-124 in constraining synaptic plasticity through CREB
Abstract
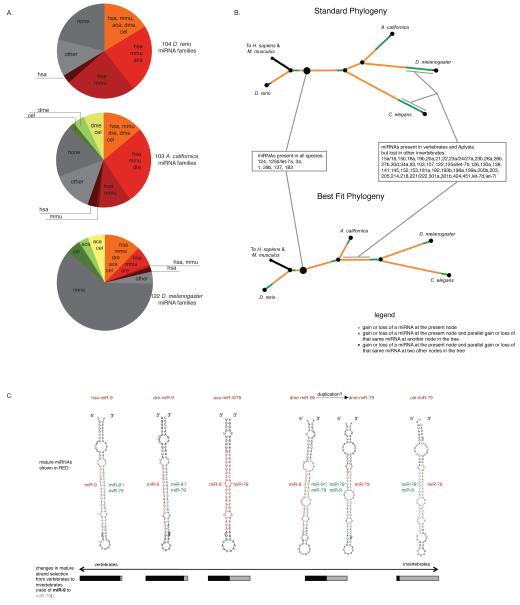
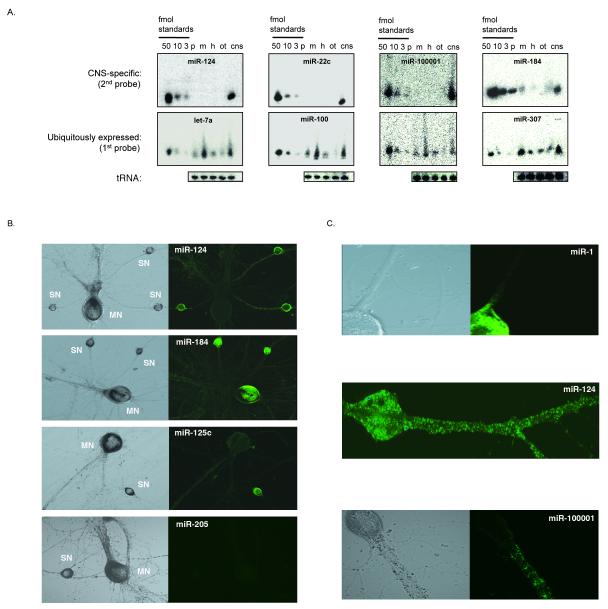
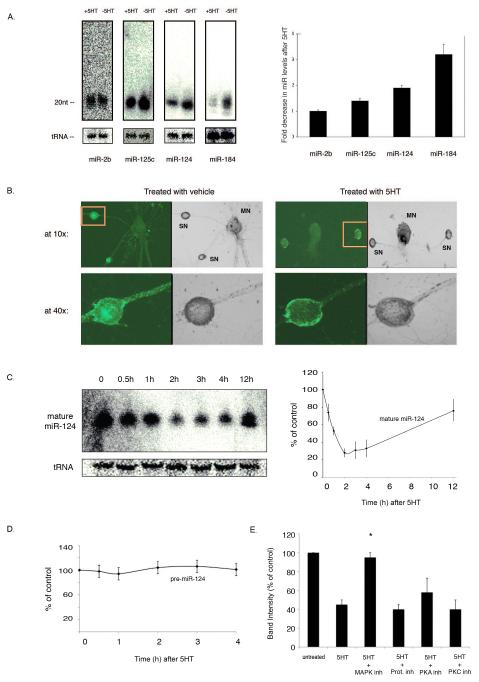
Memory storage and memory-related synaptic plasticity rely on precise spatiotemporal regulation of gene expression. To explore the role of small regulatory RNAs in learning-related synaptic plasticity, we carried out massive parallel sequencing to profile the small RNAs of Aplysia californica. We identified 170 distinct miRNAs, 13 of which were novel and specific to Aplysia. Nine miRNAs were brain enriched, and several of these were rapidly downregulated by transient exposure to serotonin, a modulatory neurotransmitter released during learning. Further characterization of the brain-enriched miRNAs revealed that miR-124, the most abundant and well-conserved brain-specific miRNA, was exclusively present presynaptically in a sensory-motor synapse where it constrains serotonin-induced synaptic facilitation through regulation of the transcriptional factor CREB. We therefore present direct evidence that a modulatory neurotransmitter important for learning can regulate the levels of small RNAs and present a role for miR-124 in long-term plasticity of synapses in the mature nervous system.
Figures



Comment in
-
MicroRNAs in memory processing.Neuron. 2009 Sep 24;63(6):714-6. doi: 10.1016/j.neuron.2009.09.007. Neuron. 2009. PMID: 19778498
Similar articles
-
The two regulatory subunits of aplysia cAMP-dependent protein kinase mediate distinct functions in producing synaptic plasticity.J Neurosci. 2004 Mar 10;24(10):2465-74. doi: 10.1523/JNEUROSCI.4331-03.2004. J Neurosci. 2004. PMID: 15014122 Free PMC article.
-
A transient, neuron-wide form of CREB-mediated long-term facilitation can be stabilized at specific synapses by local protein synthesis.Cell. 1999 Oct 15;99(2):221-37. doi: 10.1016/s0092-8674(00)81653-0. Cell. 1999. PMID: 10535740
-
Synapse-specific, long-term facilitation of aplysia sensory to motor synapses: a function for local protein synthesis in memory storage.Cell. 1997 Dec 26;91(7):927-38. doi: 10.1016/s0092-8674(00)80484-5. Cell. 1997. PMID: 9428516
-
Postsynaptic regulation of the development and long-term plasticity of Aplysia sensorimotor synapses in cell culture.J Neurobiol. 1994 Jun;25(6):666-93. doi: 10.1002/neu.480250608. J Neurobiol. 1994. PMID: 8071666 Review.
-
Synaptic remodeling, synaptic growth and the storage of long-term memory in Aplysia.Prog Brain Res. 2008;169:179-98. doi: 10.1016/S0079-6123(07)00010-6. Prog Brain Res. 2008. PMID: 18394474 Review.
Cited by
-
The DEAH-box helicase DHX36 mediates dendritic localization of the neuronal precursor-microRNA-134.Genes Dev. 2013 May 1;27(9):991-6. doi: 10.1101/gad.211243.112. Genes Dev. 2013. PMID: 23651854 Free PMC article.
-
The Path to microRNA Therapeutics in Psychiatric and Neurodegenerative Disorders.Front Genet. 2012 May 17;3:82. doi: 10.3389/fgene.2012.00082. eCollection 2012. Front Genet. 2012. PMID: 22629284 Free PMC article.
-
Validation of differentially expressed brain-enriched microRNAs in the plasma of PD patients.Ann Clin Transl Neurol. 2020 Sep;7(9):1594-1607. doi: 10.1002/acn3.51146. Epub 2020 Aug 29. Ann Clin Transl Neurol. 2020. PMID: 32860338 Free PMC article.
-
Pathogenetic and therapeutic applications of microRNAs in major depressive disorder.Prog Neuropsychopharmacol Biol Psychiatry. 2016 Jan 4;64:341-8. doi: 10.1016/j.pnpbp.2015.02.003. Epub 2015 Feb 14. Prog Neuropsychopharmacol Biol Psychiatry. 2016. PMID: 25689819 Free PMC article. Review.
-
Molecular and cellular pathophysiology of preclinical Alzheimer's disease.Behav Brain Res. 2016 Sep 15;311:54-69. doi: 10.1016/j.bbr.2016.05.030. Epub 2016 May 13. Behav Brain Res. 2016. PMID: 27185734 Free PMC article. Review.
References
-
- Alberini CM, Ghirardi M, Metz R, Kandel ER. C/EBP is an immediate-early gene required for the consolidation of long-term facilitation in Aplysia. Cell. 1994;76:1099–114. - PubMed
-
- Aravin AA, Lagos-Quintana M, Yalcin A, Zavolan M, Marks D, Snyder B, Gaasterland T, Meyer J, Tuschl T. The small RNA profile during Drosophila melanogaster development. Dev Cell. 2003;5:337–50. - PubMed
-
- Ashraf SI, McLoon AL, Sclarsic SM, Kunes S. Synaptic protein synthesis associated with memory is regulated by the RISC pathway in Drosophila. Cell. 2006;124:191–205. - PubMed
-
- Barco A, Alarcon JM, Kandel ER. Expression of constitutively active CREB protein facilitates the late phase of long-term potentiation by enhancing synaptic capture. Cell. 2002;108:689–703. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
