

An alternative pathway mediates the mouse and human cone visual cycle
- PMID: 19781940
- PMCID: PMC2762012
- DOI: 10.1016/j.cub.2009.07.054
An alternative pathway mediates the mouse and human cone visual cycle
Abstract
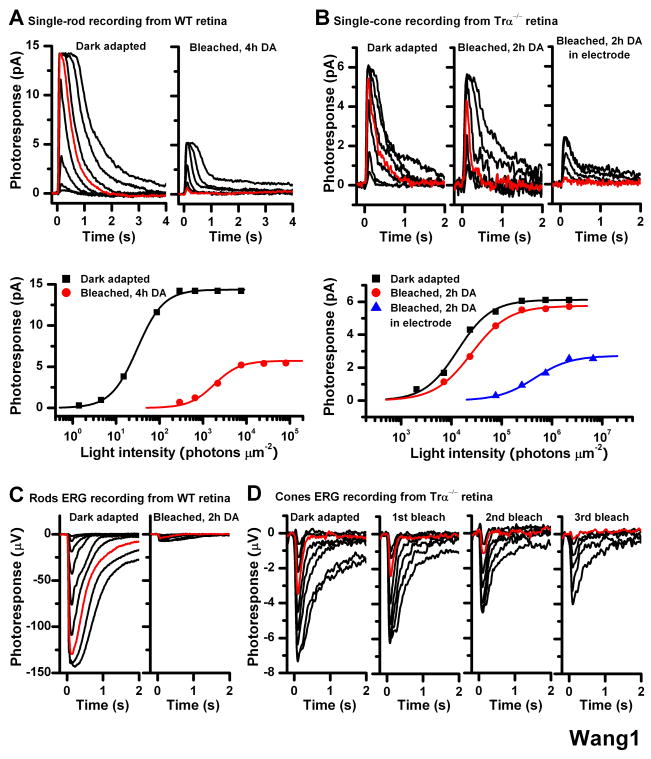
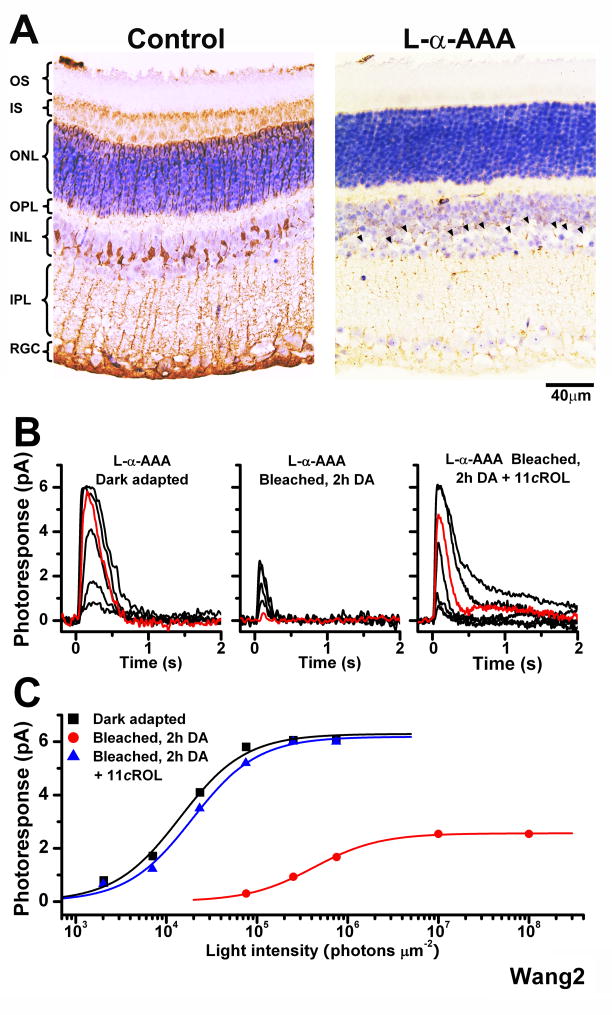
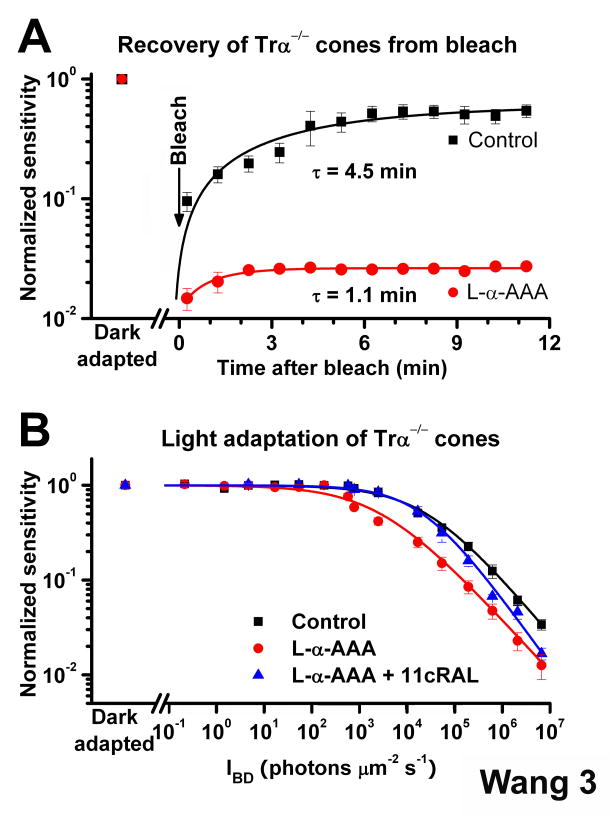
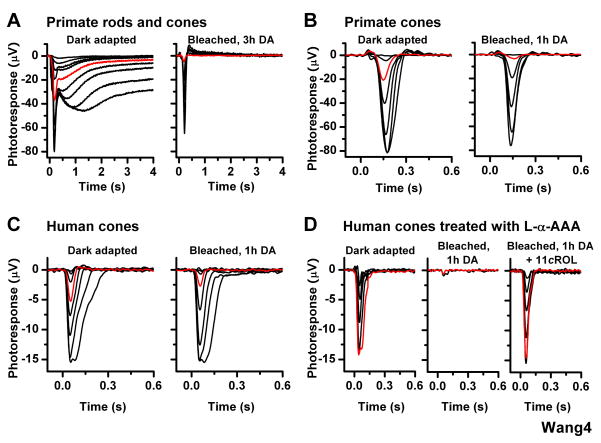
One of the fundamental mysteries of the human visual system is the continuous function of cone photoreceptors in bright daylight. As visual pigment is destroyed, or bleached, by light, cones require its rapid regeneration, which in turn involves rapid recycling of the pigment's chromophore. The canonical visual cycle for rod and cone pigments involves recycling of their chromophore from all-trans retinol to 11-cis retinal in the pigment epithelium, adjacent to photoreceptors. However, shortcomings of this pathway indicate the function of a second, cone-specific, mechanism for chromophore recycling. Indeed, biochemical and physiological studies on lower species have described a cone-specific visual cycle in addition to the long-known pigment epithelium pathway. Two important questions remain, however: what is the role of this pathway in the function of mammalian cones, and is it present in higher mammals, including humans? Here, we show that mouse, primate, and human neural retinas promote pigment regeneration and dark adaptation selectively in cones, but not in rods. This pathway supports rapid dark adaptation of mammalian cones and extends their dynamic range in background light independently of the pigment epithelium. This pigment-regeneration mechanism is essential for our daytime vision and appears to be evolutionarily conserved.
Figures
References
-
- Ebrey T, Koutalos Y. Vertebrate photoreceptors. Prog Retin Eye Res. 2001;20:49–94. - PubMed
-
- Saari JC. Biochemistry of visual pigment regeneration: the Friedenwald lecture. Invest Ophthalmol Vis Sci. 2000;41:337–348. - PubMed
-
- Bustamante JJ, Ziari S, Ramirez RD, Tsin AT. Retinyl ester hydrolase and the visual cycle in the chicken eye. Am J Physiol. 1995;269:R1346–1350. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
