

Review
doi: 10.1016/j.molcel.2009.09.005.
Transcription dynamics
Affiliations
- PMID: 19782025
- PMCID: PMC6326382
- DOI: 10.1016/j.molcel.2009.09.005
Item in Clipboard
Review
Transcription dynamics
Mol Cell.
.
Abstract
All aspects of transcription and its regulation involve dynamic events. The basal transcription machinery and regulatory components are dynamically recruited to their target genes, and dynamic interactions of transcription factors with chromatin--and with each other--play a key role in RNA polymerase assembly, initiation, and elongation. These short-term binding dynamics of transcription factors are superimposed by long-term cyclical behavior of chromatin opening and transcription factor-binding events. Its dynamic nature is not only a fundamental property of the transcription machinery, but it is emerging as an important modulator of physiological processes, particularly in differentiation and development.
Figures
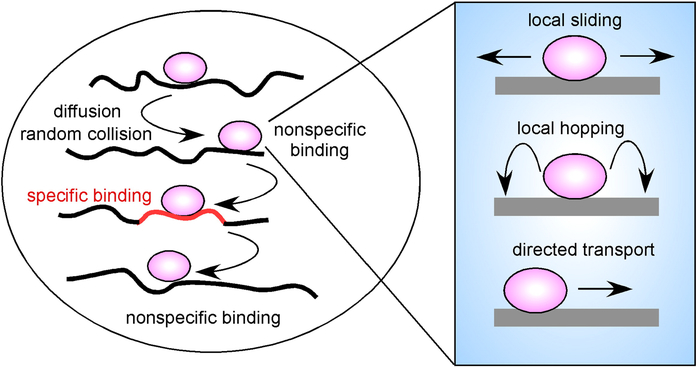
A TF (purple) diffuses through the nuclear space and by random collision associates with chromatin. Most encounters are at nontarget sites resulting in highly transient interactions. Occasionally, a specific binding site (orange) is encountered, and prolonged binding occurs. At each encounter a TF might undergo local motion on the chromatin fiber by either sliding along the DNA, hopping locally or by directed, motor driven motion.
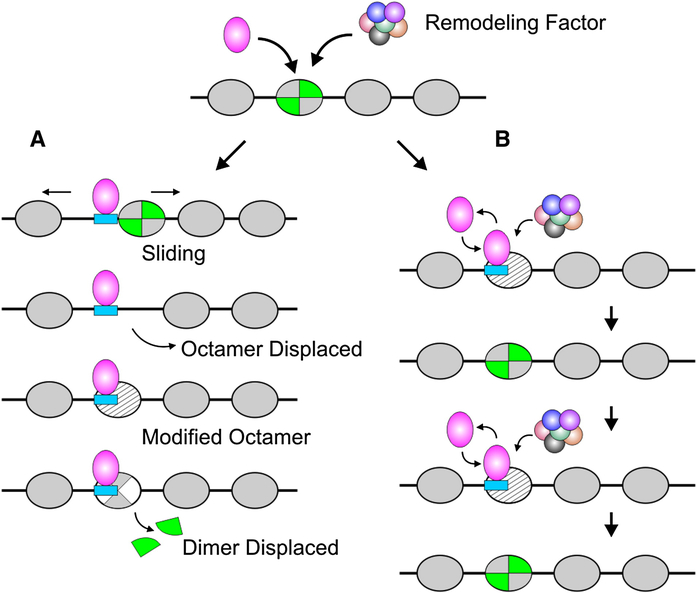
(A) Local nucleosome reorganization, giving rise to enhanced transcription factor access, is commonly discussed in terms of altered static states. Evidence has been advanced to support several specific mechanisms, including sliding to a new position, octamer displacement, modified octamer structure, and partial octamer dissociation. These modified nucleosome states would in turn accommodate factor binding events not compatible with the unaltered state. (B) A dynamic view of local transitions suggests that remodeling is a continuous process. Remodeling complexes are targeted to specific nucleosomes by a given transcription factor. However, both the remodeling process itself and commensurate binding of a factor are transient events. Constant repetition of this cycle produces a shift in the equilibrium distribution of both the transcription factor and nucleosome components.
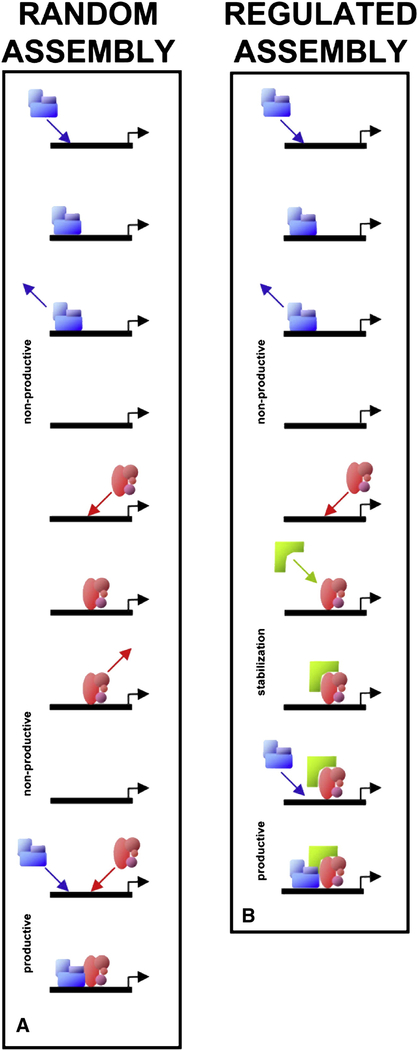
(A) Two subcomplexes (blue and red) exhibit dynamic exchange at a promoter template (black line). Assembly of the full complex occurs via random collisions with the template, leading sometimes to nonproductive interactions and occasionally to productive interactions (the simultaneous presence of both subcomplexes). (B) Regulation of dynamic exchange rates can facilitate or inhibit complex formation. Here, binding of the red subcomplex is stabilized by the bracket-shaped molecule (green), leading to a longer dwell time on the promoter template, thereby facilitating assembly of the complex. Alternatively, destabilization of factor binding could also occur, inhibiting assembly (not shown).
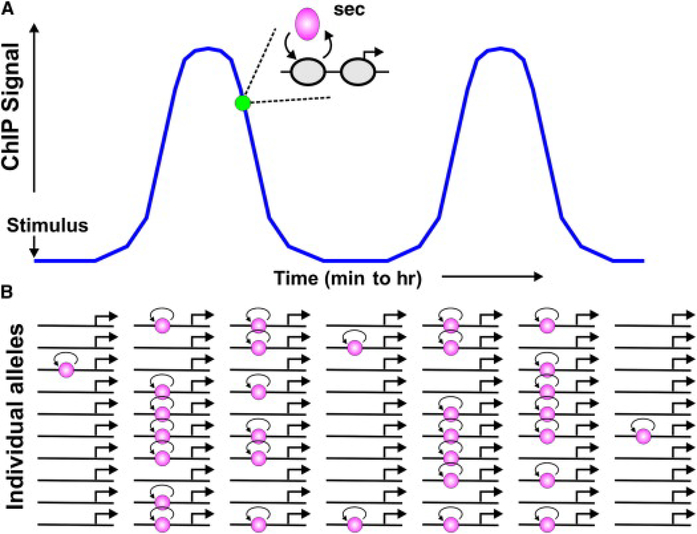
(A) Most factors that have been studied in living cells exchange rapidly, on a time scale of seconds, with their recognition elements in chromatin (“hit-and-run”). The frequency and transient duration of these binding events can also fluctuate on a longer time scale, by a variety of mechanisms. (B) Depicted here are a set of ten abstracted alleles with one binding element. After a transcriptional stimulus, the number of interaction events increases. If this element is sampled across the population by a methodology such as ChIP, more of the events will be captured in a given time. If secondary mechanisms are triggered that decrease the interaction frequency, the ChIP signal will decrease, and an oscillatory process may ensue. However, if real-time residence times could be examined at a specific allele (green circle), one would observe rapid exchange.
References
-
- Araki R, Fukumura R, Sasaki N, Kasama Y, Suzuki N, Takahashi H, Tabata Y, Saito T, and Abe M (2006). More than 40,000 transcripts, including novel and noncoding transcripts, in mouse embryonic stem cells. Stem Cells 24, 2522–2528. - PubMed
-
- Azuara V, Perry P, Sauer S, Spivakov M, Jorgensen HF, John RM, Gouti M, Casanova M, Warnes G, Merkenschlager M, and Fisher AG (2006). Chromatin signatures of pluripotent cell lines. Nat. Cell Biol 8, 532–538. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
