

Silymarin inhibits in vitro T-cell proliferation and cytokine production in hepatitis C virus infection
- PMID: 19782083
- PMCID: PMC2819600
- DOI: 10.1053/j.gastro.2009.09.021
Silymarin inhibits in vitro T-cell proliferation and cytokine production in hepatitis C virus infection
Abstract
Background & aims: Silymarin, an extract from the seeds of the milk thistle plant Silybum marianum, has been used for centuries for the treatment of chronic liver diseases. Despite common use by patients with hepatitis C in the United States, its clinical efficacy remains uncertain. The goal of this study was to determine whether silymarin has in vitro effects on immune function that might have implications for its potential effect on hepatitis C virus (HCV)-induced liver disease.
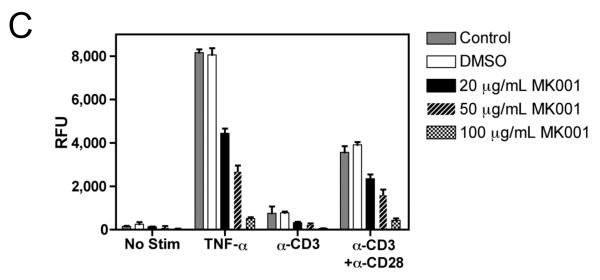
Methods: Freshly isolated peripheral blood mononuclear cells (PBMC) and T cells from HCV-infected and uninfected subjects were tested in vitro for responses to nonspecific and antigenic stimulation in the presence and absence of a standardized preparation of silymarin (MK001).
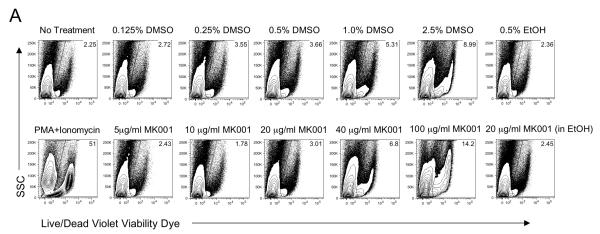
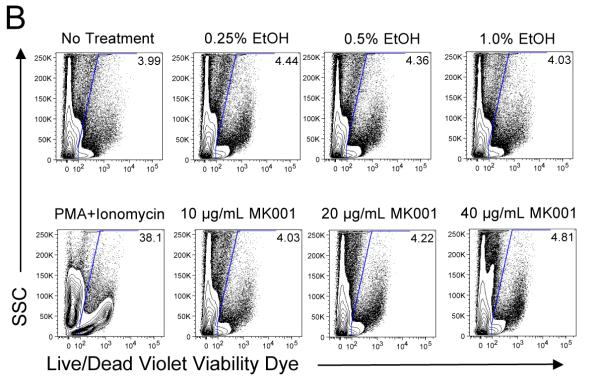
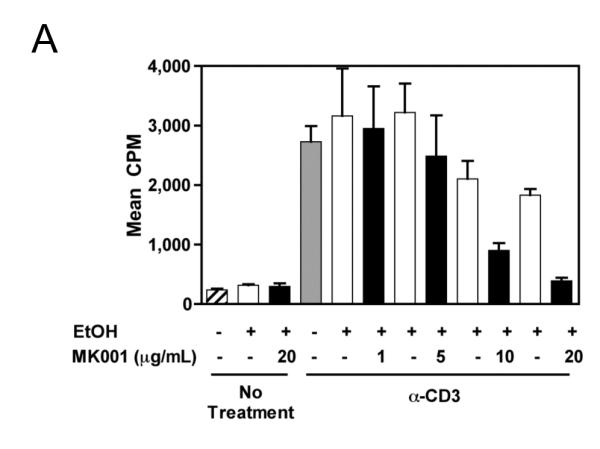
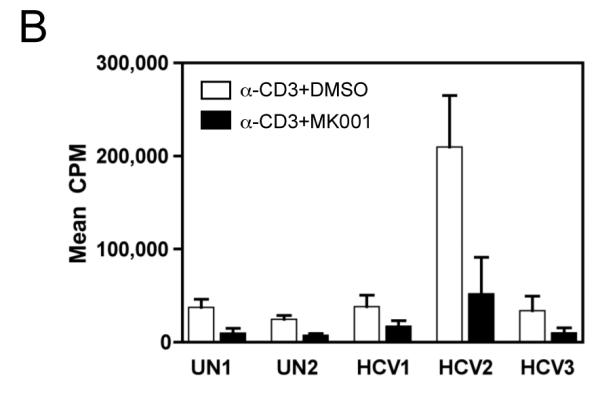
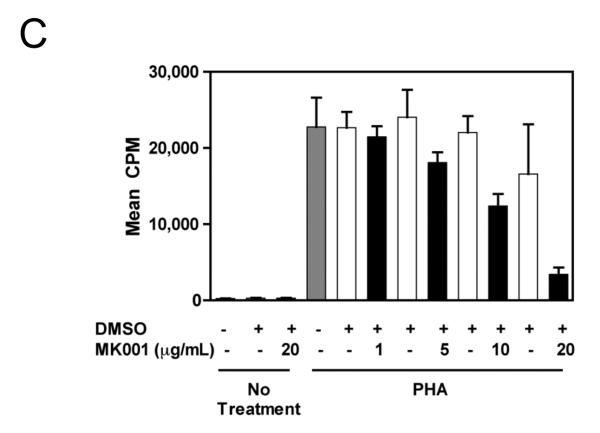
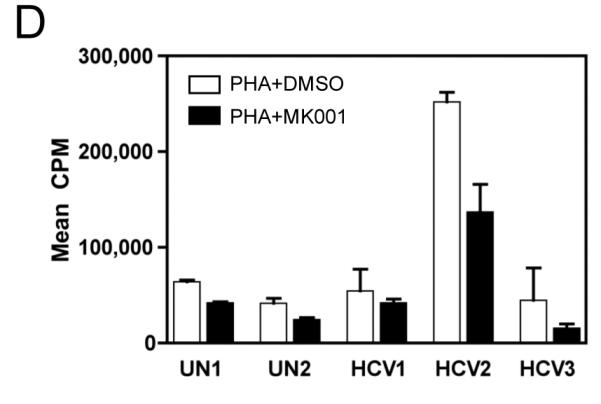
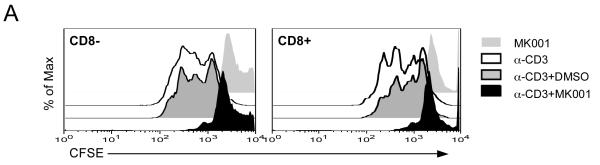
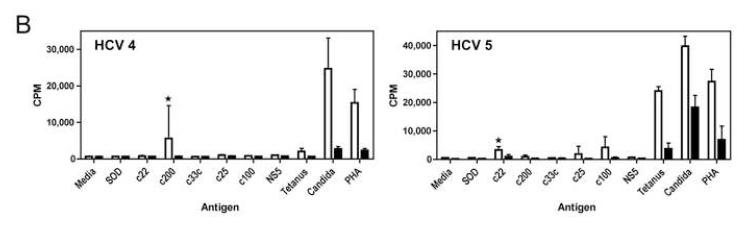
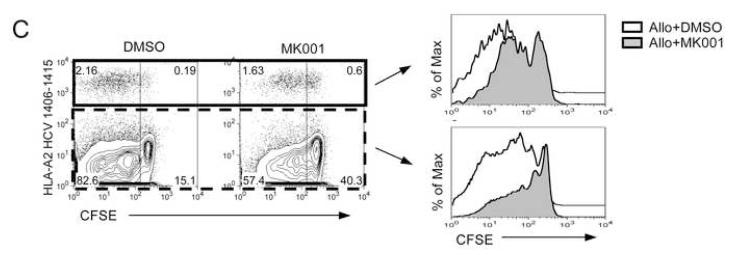
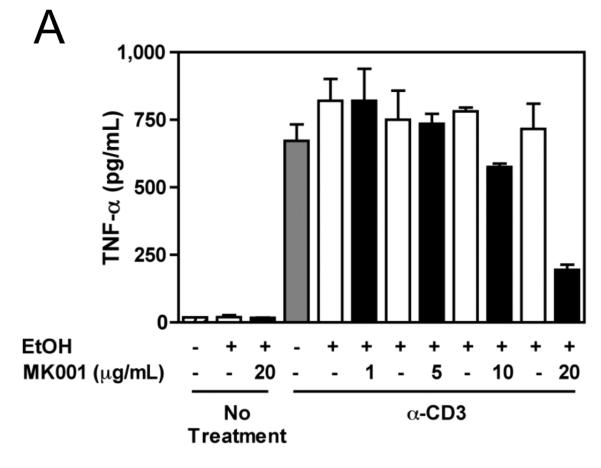
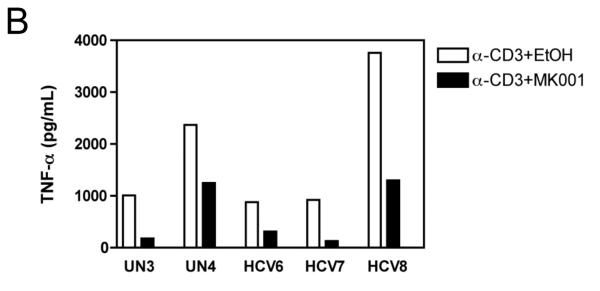
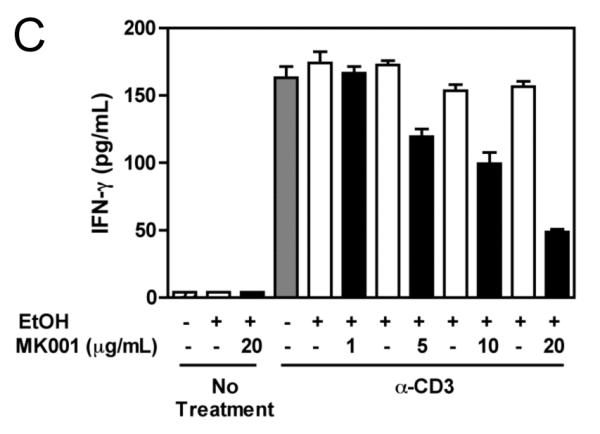
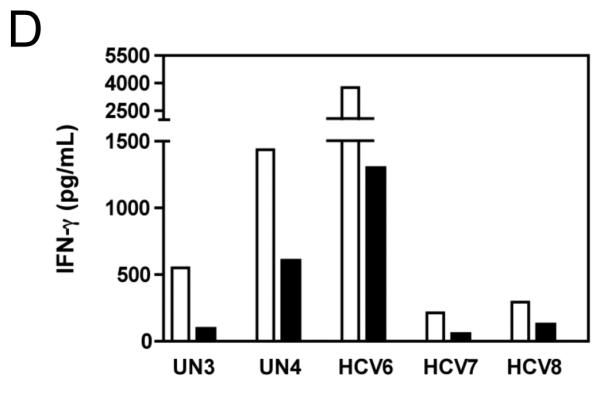
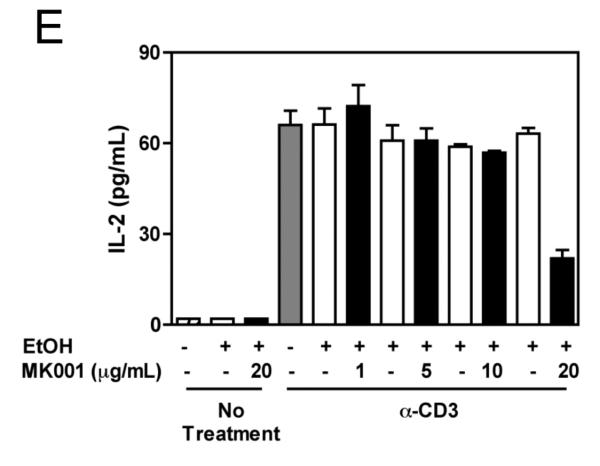
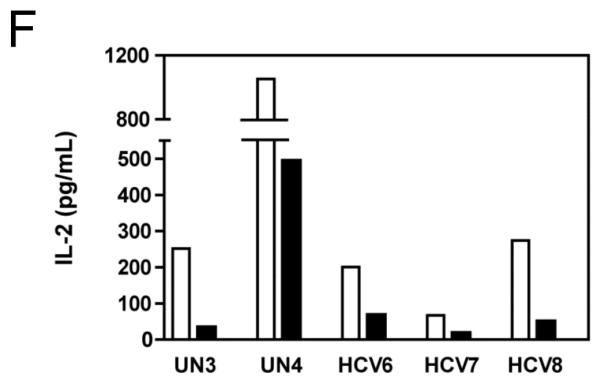
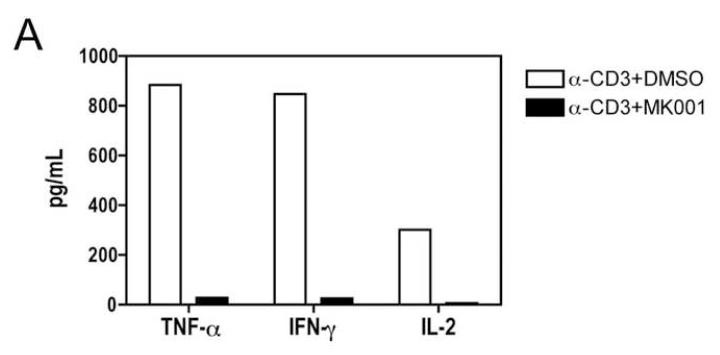
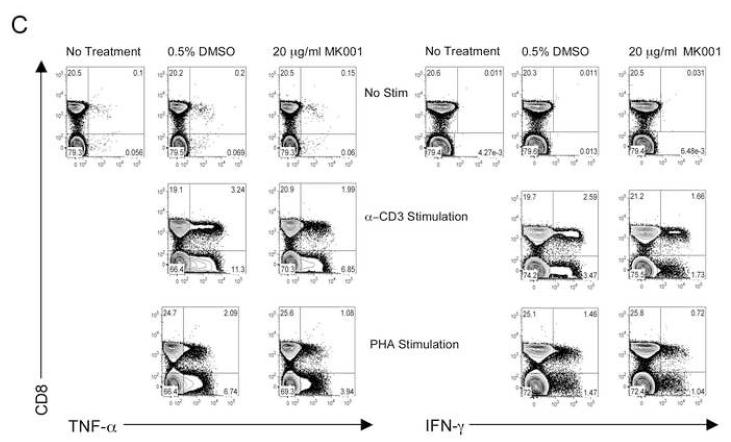
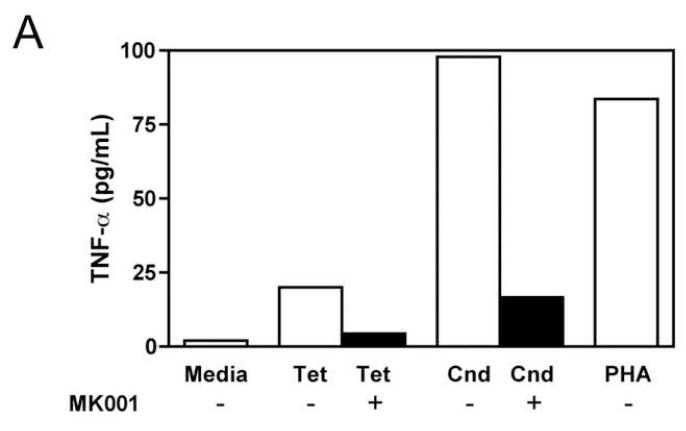
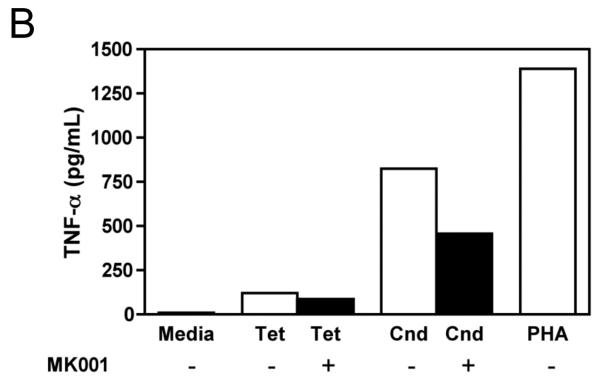
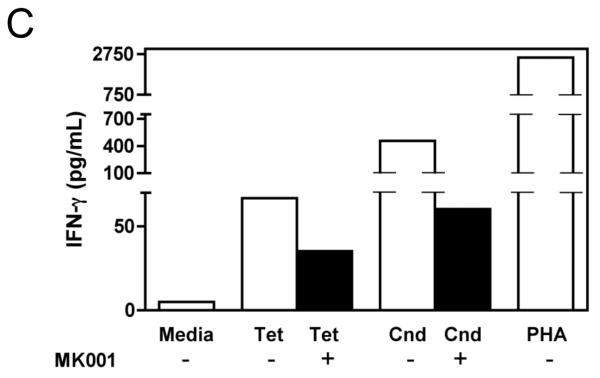
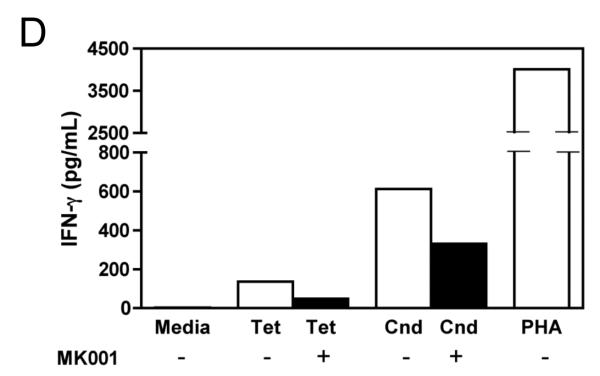
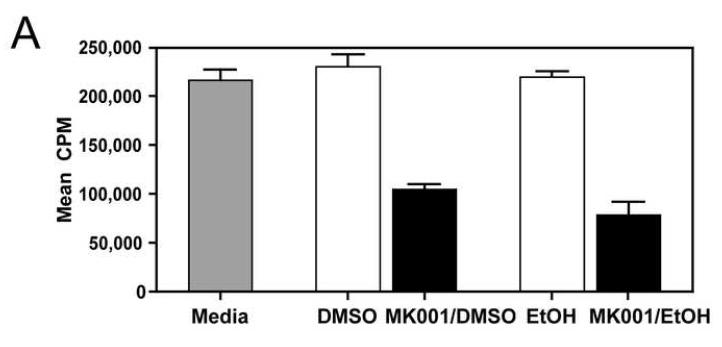
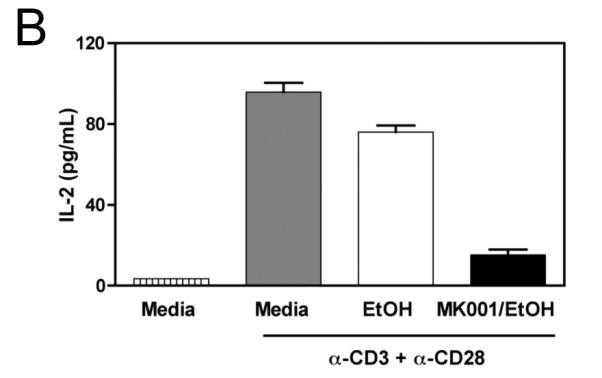
Results: Minimal MK001 toxicity on PBMC was found at concentrations between 5 and 40 microg/mL. MK001 dose dependently inhibited the proliferation and secretion of tumor necrosis factor (TNF)-alpha, interferon (IFN)-gamma, and interleukin (IL)-2 by PBMC stimulated with anti-CD3. In addition, MK001 inhibited proliferation by CD4(+) T cells to HCV, Candida, and tetanus protein antigens and by HLA-A2/HCV 1406-1415-specific CD8(+) T cells to allogeneic stimulation. MK001 inhibited T-cell TNF-alpha and IFN-gamma cytokine secretion to tetanus and Candida protein antigens. Finally, MK001 inhibited nuclear factor-kappaB transcriptional activation after T-cell receptor-mediated stimulation of Jurkat T cells, consistent with its ability to inhibit Jurkat T-cell proliferation and secretion of IL-2.
Conclusions: Silymarin's ability to inhibit the proliferation and proinflammatory cytokine secretion of T cells, combined with its previously described antiviral effect, suggests a possible mechanism of action that could lead to clinical benefit during HCV infection.
Figures
References
-
- Alter HJ, Seeff LB. Recovery, persistence, and sequelae in hepatitis C virus infection: a perspective on long-term outcome. Seminars in Liver Disease. 2000;20:17–35. - PubMed
-
- Manns MP, McHutchison JG, Gordon SC, Rustgi VK, Shiffman M, Reindollar R, Goodman ZD, Koury K, Ling M, Albrecht JK. Peginterferon alfa-2b plus ribavirin compared with interferon alfa-2b plus ribavirin for initial treatment of chronic hepatitis C: a randomised trial. Lancet. 2001;358:958–65. - PubMed
-
- Fried MW, Shiffman ML, Reddy KR, Smith C, Marinos G, Goncales FL, Haussinger D, M D, G C, Dhumeaux D, Craxi A, Lin A, Hoffmann J, Yu J. Peginterferon alfa-2a plus ribavirin for chronic hepatitis C virus infection. New England Journal of Medicine. 2002;347:975–982. - PubMed
-
- Davis GL, Albright JE, Cook SF, Rosenberg DM. Projecting future complications of chronic hepatitis C in the United States. Liver Transpl. 2003;9:331–8. - PubMed
-
- Strader DB, Bacon BR, Lindsay KL, La Brecque DR, Morgan T, Wright EC, Allen J, Khokar MF, Hoofnagle JH, Seeff LB. Use of complementary and alternative medicine in patients with liver disease. Am J Gastroenterol. 2002;97:2391–7. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
