

Quantitative assignment of reaction directionality in constraint-based models of metabolism: application to Escherichia coli
- PMID: 19783351
- PMCID: PMC2802319
- DOI: 10.1016/j.bpc.2009.08.007
Quantitative assignment of reaction directionality in constraint-based models of metabolism: application to Escherichia coli
Abstract
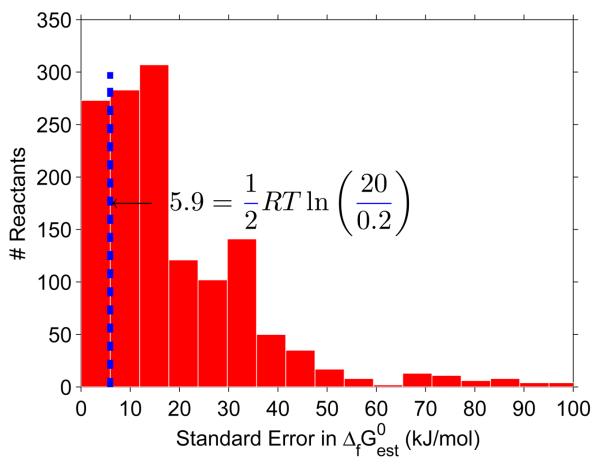
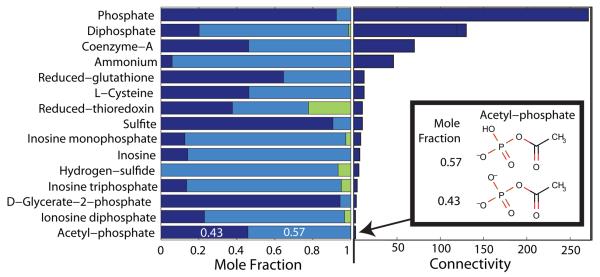
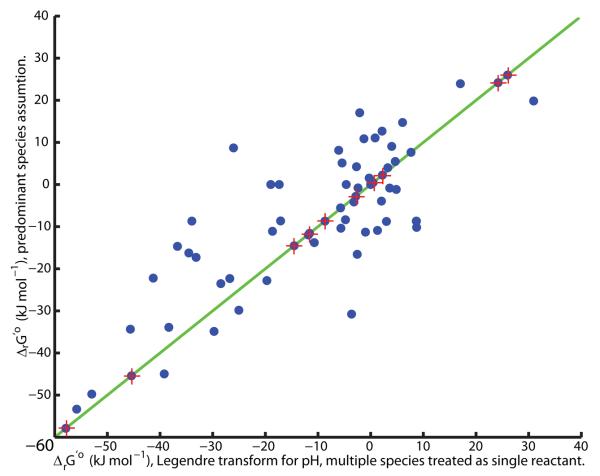
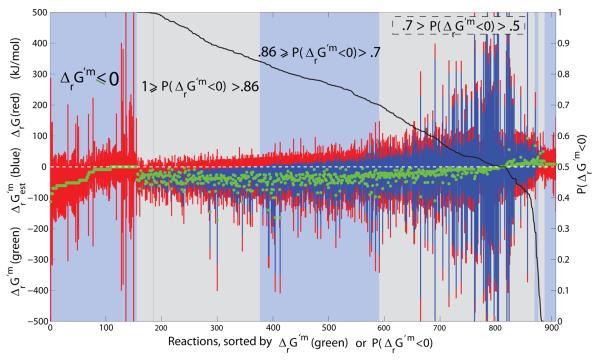
Constraint-based modeling is an approach for quantitative prediction of net reaction flux in genome-scale biochemical networks. In vivo, the second law of thermodynamics requires that net macroscopic flux be forward, when the transformed reaction Gibbs energy is negative. We calculate the latter by using (i) group contribution estimates of metabolite species Gibbs energy, combined with (ii) experimentally measured equilibrium constants. In an application to a genome-scale stoichiometric model of Escherichia coli metabolism, iAF1260, we demonstrate that quantitative prediction of reaction directionality is increased in scope and accuracy by integration of both data sources, transformed appropriately to in vivo pH, temperature and ionic strength. Comparison of quantitative versus qualitative assignment of reaction directionality in iAF1260, assuming an accommodating reactant concentration range of 0.02-20mM, revealed that quantitative assignment leads to a low false positive, but high false negative, prediction of effectively irreversible reactions. The latter is partly due to the uncertainty associated with group contribution estimates. We also uncovered evidence that the high intracellular concentration of glutamate in E. coli may be essential to direct otherwise thermodynamically unfavorable essential reactions, such as the leucine transaminase reaction, in an anabolic direction.
Figures
References
-
- Palsson BØ. Systems Biology: Properties of Reconstructed Networks. Cambridge University Press; Cambridge: 2006.
-
- Pál C, Papp B, Lercher MJ, Csermely P, Oliver SG, Hurst LD. Chance and necessity in the evolution of minimal metabolic networks. Nature. 2006;440(7084):667–670. - PubMed
-
- Mahadevan R, Schilling CH. The effects of alternate optimal solutions in constraint-based genome-scale metabolic models. Metab Eng. 2003;5(4):264–276. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
